
Sex Differences in Exploratory Behaviour of Laboratory CD-1 Mice (Mus musculus)
by Toyohito Tanaka
Division of Toxicology, Tokyo Metropolitan Institute of Public
Health
 Correspondence: Toyohito Tanaka
Correspondence: Toyohito Tanaka
Division of Toxicology, Tokyo Metropolitan Institute of Public Health,
3-24-1, Hyakunincho, Shinjuku-ku, Tokyo 169-0073, Japan
Tel +81-3-3363-3231 ex. 5603
Fax +81-3-3368-4060
E-mail t-tanaka@poohlover.net
Summary
Sex differences in exploratory behaviour were investigated in 120 litters of control CD-1 mice in two-generation toxicity studies over a 13-year period. Exploratory behaviour of F1-generation mice was investigated at 3 weeks of age during the lactation period and at 8 weeks of age after weaning. In the F1 generation, at 3 weeks of age, female offspring showed less activity in terms of the number of movements, movement time, number of vertical activities and vertical time than male offspring. Longitudinal analysis showed that these variables were parallel in males and females during the 10-minute observation period. At 8 weeks of age, female offspring in the F1 generation showed less activity with respect to the number of movements, movement time, average distance, number of vertical activities and vertical time but showed more horizontal activities than male offspring. The number of movements, movement time and vertical time were not parallel in longitudinal analysis; however, all other variables were parallel. Sex differences in exploratory behaviour of immature mice showed quantitative changes only. In adult mice, variables for rearing excluding vertical time showed quantitative changes only; however, variables for ambulation and vertical time showed quantitative and qualitative changes. From these results, it appears that female CD-1 mice display less exploratory behaviour than male CD-1 mice and that these sex differences in exploratory behaviour occur from the prepubertal period.
Introduction
Several studies have reported differences in nonsexual behaviour between male and female mammals. In particular, differences in emotional behaviour have been frequently observed in rodents. Valle(1970) reported that female rats were generally more active than male rats in the open field. Masur(1972) found that male rats displayed more defecation and less ambulatory behaviour than female rats in the open field. Archer(1974, 1977) reported a lower latency time and more stationary sniffing among male rats and mice compared with those in female rats and mice. Archer(1975) suggested that male rats typically showed more defecation behaviour than female rats in the open field. However, among mice, neither sex can be said to typically defecate more than the other. In general, in both rats and mice, males show fewer ambulatory movements than females in open-field tests; however, there have also been several cases where no significant between-sex differences have been observed. Gray (1979) suggested that similar to outbred rat strains, outbred mice strains exhibit the typical rat pattern of greater defecation and less activity among males. Nevertheless, few studies have examined sex differences in exploratory behaviour of outbred mice.
In reproductive and developmental toxicity studies, exploratory behaviour is usually measured as a neurobehavioural parameter, and it is an important variable for detecting neurobehavioural toxicity. At the Tokyo Metropolitan Institute of Public Health, several reproductive and neurobehavioural toxicity studies of food additives have been performed on CD-1 (ICR) mice (Mus musculus), and their results have been evaluated. Exploratory behaviour, motor activity in a novel environment including ambulatory and rearing movements, has been analysed in immature and adult F1-generation mice. Chemical effects at times result in different patterns of exploratory behaviour in both sexes. However, there have been very few reports on exploratory behaviour of control mice, in particular, in terms of sex-related differences. This study deals with sex differences in exploratory behaviour of control CD-1 (ICR) mice in two-generation toxicity studies under similar conditions over a 13-year-period.
Materials and methods
Animals and maintenance
Male and female mice (Crj: CD-1, 4 weeks old) were purchased from
Charles River Japan Inc., Kanagawa, Japan, for each study over a
13-year-period. They were housed individually in polycarbonate
solid-floored cages with wood flakes (prepared from American fir tree;
Nihon Clea, Tokyo, Japan) and were kept in a temperature-controlled
room maintained at 24°C ± 1°C with a relative humidity of 55% ± 5%
under a 12-h light/12-h dark cycle. They were given a controlled
basal diet (CE-2; Nihon Clea) and water ad libitum.
The present study was conducted under guidelines set by theNational Research Council (1996) and the Science Council of Japan (2006). Animal experiments conformed with the following Japanese laws and relevant regulations: ‘Act on Welfare and Management of Animals’ (Act No. 105 of 1 October 1973, revised on 22 June 2005); Notice No. 88 of 28 April 2006 of the Ministry of the Environment of Japan ‘Standards Relating to the Care and Management of Laboratory Animals and Relief of Pain’; and Notification of 1 June 2006 of the Ministry of Health, Labour and Welfare of Japan ‘Fundamental Guidelines for Proper Conduct of Animal Experiments and Related Activities in Research Institutions under the jurisdiction of the Ministry of Health, Labour and Welfare’.
Reproductive procedure
In 14 reproductive and neurobehavioural toxicity studies (1993–2005),
male and female mice (n = 10 per study) were used as a control group.
F0 generation mice were 5 weeks old at the start of the study. At 9
weeks of age, each female mouse was paired with a male from the
control group. After 5 days, the males were removed and the females
were allowed to carry their litters to term, deliver and rear their
offspring. Offspring were weaned at 4 weeks of age, and one male and
one female mouse were randomly selected from each litter for the
remaining experiment. They were housed individually in polycarbonate
solid-floored cages with wood flakes after weaning. These procedures
were the same in each study. A total of 1,594 offspring were born in
126 litters. After the lactation period, 120 litters were weaned at 4
weeks of age.
Exploratory behaviour
Exploratory behaviour of the mice was measured using an animal
movement analyzing system (ANIMATE AT-420; Toyo Sangyo Co., Ltd.,
Toyama, Japan) at 3 and 8 weeks of age in the F1 generation. This
system consisted of a doughnut-shaped cage made from acrylate resins
with 36 near-infrared photosensors used to measure spontaneous motor
activity (Matsumoto et al., 1990a, b). Behavioural
variables were recorded for 10 min at 1-min intervals for one male and
one female randomly selected from each litter at 3 weeks of age and
for all animals (one male and one female per litter) at 8 weeks of age
in the F1 generation. These procedures were the same in each study.
The measured variables were as detailed below (Tanaka,
2010):
-
Number of movements: Frequency of changes in location
information.
-
Movement time (s): Total time for which the animal was moving.
-
Number of horizontal activities: Frequency at which the animal began
moving from a stationary position.
-
Total distance (cm): Total distance covered by the animal after it
moved from the central position.
-
Number of vertical activities: Frequency of rearing (including
jumping).
- Vertical time (s): Total rearing time.
-
Number of turns: Frequency at which the direction of movement
changed by 45° or more.
-
Average distance (cm): (total distance/number of horizontal
activities).
- Average speed (cm/s): (total distance/movement time).
- Number of defecations: number of faecal pellets.
-
Average movement time (s): (movement time/number of horizontal
activities).
- Average rearing time (s): (vertical time/number of vertical activities).
Statistical analysis
Movement activity data related to exploratory behaviour of male and
female mice were compared using the Wilcoxon test (Martin & Bateson, 1990). Differences in the longitudinal patterns (during
the 10-minute observation period) of exploratory behaviour between
sexes were assessed using profile analysis (test for equality of mean
vectors, test for equality of covariance matrices and the parallelism
hypothesis test). This longitudinal analysis is appropriate for a
study that involves repeated observations of the same variables over
long periods of time (Fujikoshi et al., 2008).
Differences between sexes at different time points were assessed using
the Steel–Dwass test (Martin & Bateson, 1990).
Results
At 3 weeks of age, exploratory behaviour of 112 male and 111 female offspring was examined. Female offspring showed less activity than male offspring in terms of the following variables: number of movements, movement time (s), number of vertical activities and vertical time (s) (Table 1). These variables were parallel in longitudinal analysis in male and female offspring (Table 2), and the distances between the lines were significantly different (P < 0.001, 0.05, 0.05 and 0.05, respectively). The number of movements was significantly different at 1, 2 and 4 min between male and female offspring (Fig. 1). The other variables were not significantly different between male and female offspring (Table 1).
Table 1. Summary of differences in exploratory behaviour between male and female F1-generation mice during the 10-minute observation period.
| Male | Female | |
| At 3 weeks of age | ||
| No. of mice | 112 | 111,00 |
| No. of movements c | 1357.5 ± 325.0 | 1173.7 ± 290.8 |
| Movement time (s) a | 438.54 ± 72.85 | 419.20 ± 73.43 |
| No. of horizontal activities | 49.5 ± 13.1 | 49.5 ± 12.6 |
| Total distance (cm) | 2220.52 ± 759.49 | 2135.63 ± 761.78 |
| Average distance (cm) | 51.22 ± 31.08 | 47.98 ± 27.02 |
| Average speed (cm/s) | 4.96 ± 1.10 | 4.99 ± 1.08 |
| No. of vertical activities a | 145.7 ± 40.6 | 133.5 ± 34.0 |
| Vertical time (s) b | 112.18 ± 35.77 | 99.69 ± 34.87 |
| No. of turnings | 50.6 ± 19.8 | 47.2 ± 17.0 |
| No. of defecations | 6.55 ± 2.50 | 6.59 ± 2.47 |
| Average movement time (s) | 9.67 ± 3.78 | 9.08 ± 3.14 |
| Average rearing time (s) | 0.78 ± 0.20 | 0.75 ± 0.21 |
| At 8 weeks of age | ||
| No. of mice | 113 | 112 |
| No. of movements c | 2147.6 ± 265.5 | 1787.8 ± 324.3 |
| Movement time (s) c | 508.02 ± 39.89 | 484.80 ± 56.41 |
| No. of horizontal activities c | 48.6 ± 11.60 | 55.7 ± 14.78 |
| Total distance (cm) | 2801.36 ± 609.77 | 2717.99 ± 1051.99 |
| Average distance (cm) b | 65.59 ± 47.48 | 61.68 ± 65.56 |
| Average speed (cm/s) | 5.48 ± 0.93 | 5.50 ± 1.61 |
| No. of vertical activities a | 149.6 ± 28.3 | 141.4 ± 27.4 |
| Vertical time (s) c | 228.79 ± 40.57 | 180.68 ± 36.50 |
| No. of turnings | 54.1 ± 14.6 | 52.5 ± 17.5 |
| No. of defecations | 5.51 ± 3.19 | 5.37 ± 3.45 |
| Average movement time (s) c | 11.42 ± 5.17 | 9.88 ± 5.28 |
| Average rearing time (s) c | 1.55 ± 0.25 | 1.30 ± 0.25 |
Each value represents the mean ± SD. Significant differences between male and female offspring: aP < 0.05, bP < 0.01, cP < 0.001.
|
|
Figure 1. Longitudinal pattern of sex differences in exploratory behaviour at 3 weeks of age in F1-generation mice. Each value represents the mean ± SE. Significant differences between sexes at different time points: aP < 0.05.
Click image to enlarge |
|
|
Figure 2. GC-MS-SIM of the heroin metabolites from the urine samples of rats addicted to morphine (a) and heroin (b). A: morphine, B: 6-AM
Click image to enlarge |
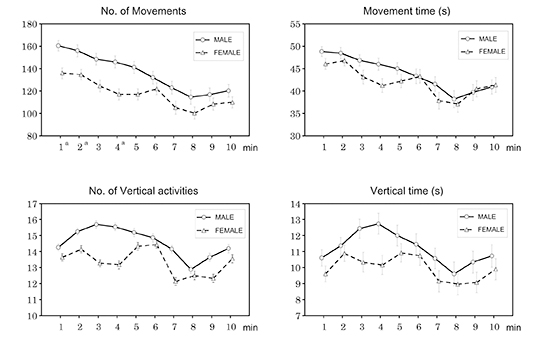
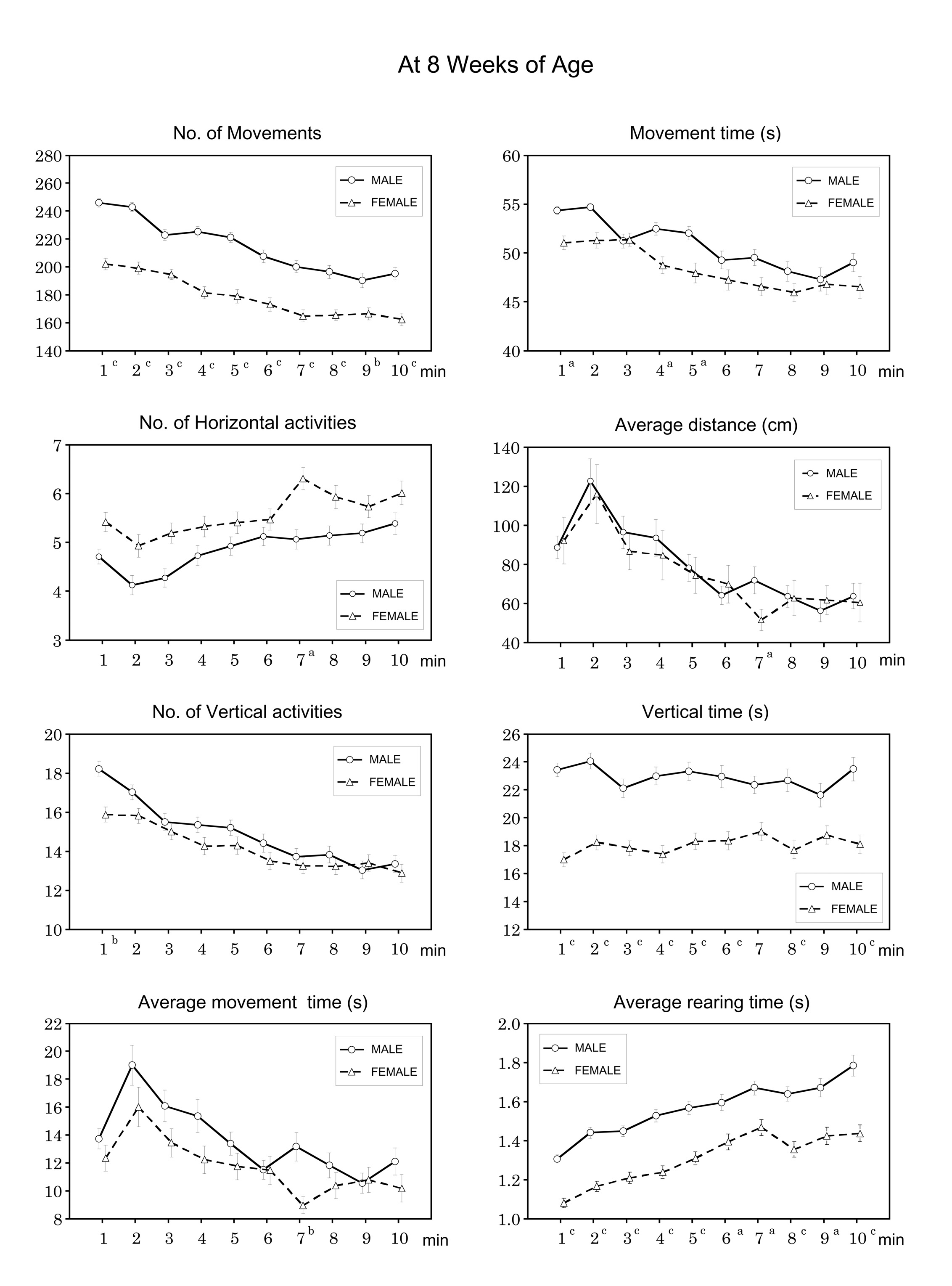
Table 2. Summary comparing longitudinal profile analysis of exploratory behaviour in male and female F1-generation mice during the 10-minute observation period.
| P-value | ||||
| Equality of mean vectors | Equality of covariance | Parallelism | Distance between lines | |
| At 3 weeks of age | ||||
| No. of movements | < 0.001 | 0.218 | 0.471 | < 0.001 |
| Movement time (s) | < 0.001 | 0.401 | 0.527 | 0.029 |
| No. of horizontal activities | < 0.001 | 0.260 | 0.117 | 0.953 |
| Total distance (cm) | < 0.001 | 0.775 | 0.216 | 0.612 |
| Average distance (cm) | < 0.001 | < 0.001 | 0.233 | 0.768 |
| Average speed (cm/s) | < 0.001 | 0.051 | 0.138 | 0.797 |
| No. of vertical activities | < 0.001 | 0.898 | 0.301 | 0.030 |
| Vertical time (s) | < 0.001 | 0.127 | 0.559 | 0.015 |
| No. of turnings | < 0.001 | 0.012 | 0.486 | 0.183 |
| Average movement time (s) | < 0.001 | < 0.001 | 0.411 | 0.246 |
| Average rearing time (s) | < 0.001 | 0.0022 | 0.831 | 0.484 |
| At 8 weeks of age | ||||
| No. of movements | < 0.001 | 0.0075 | 0.012 | < 0.001 |
| Movement time (s) | < 0.001 | < 0.001 | 0.012 | < 0.001 |
| No. of horizontal activities | < 0.001 | 0.354 | 0.457 | < 0.001 |
| Total distance (cm) | < 0.001 | < 0.001 | 0.561 | 0.700 |
| Average distance (cm) | < 0.001 | < 0.001 | 0.113 | 0.602 |
| Average speed (cm/s) | < 0.001 | < 0.001 | 0.943 | 0.848 |
| No. of vertical activities | < 0.001 | 0.534 | 0.063 | 0.023 |
| Vertical time (s) | < 0.001 | 0.0055 | 0.041 | < 0.001 |
| No. of turnings | < 0.001 | 0.061 | 0.382 | 0.393 |
| Average movement time (s) | < 0.001 | < 0.001 | 0.031 | 0.064 |
| Average rearing time (s) | < 0.001 | 0.067 | 0.428 | < 0.001 |
At 8 weeks of age, exploratory behaviour of 113 males and 112 females was examined. Females showed less activity than males in terms of the following variables: number of movements, movement time (s), average distance (cm), number of vertical activities and vertical time (s). These observations were similar to those at 3 weeks of age (Table 1). Females displayed more horizontal activities than males (Table 1); however, the average movement time (s) and average rearing time (s) were less in females than in males (Table 1).
At 8 weeks of age, the number of movements, movement time (s) and
vertical time (s) were not parallel (different longitudinal pattern
with qualitative change) according to longitudinal analysis in males
and females (Table 2); however, all other variables were parallel
(similar longitudinal pattern without qualitative changes), and the
distances between the two lines were significantly different for the
number of vertical activities, vertical time, number of horizontal
activities, movement time, number of movements (P < 0.05
or 0.001). The average movement time (s) was not parallel in
longitudinal analysis (Table 2); however, the average rearing time was
parallel, and the distance between the two lines was statistically
significant (P < 0.001).
At 8 weeks of age, the number of movements was significantly different
between male and female mice during the study period (Fig. 2). The
movement time (s) was significantly different between males and
females at 1, 4 and 5 min. The number of horizontal activities and
average distance (cm) were significantly different between the sexes
at 7 min. The number of vertical activities was significantly
different between the sexes at 1 min. Between-sex differences in
vertical time (s) were significant throughout the measuring period,
except at 7 and 9 min. The average movement time (s) was
significantly different between the sexes at 7 min (Fig. 2), and the
average rearing time (s) was also different between the sexes at all
time points. No significant difference between males and females was
observed for any other variable (Table 1).
The dispersion (extent of variation) of most variables, except for number of vertical activities, was similar between the sexes at 3 weeks of age (Table 1). At 8 weeks of age, the dispersion of most variables was greater in females than in males, in particular, in terms of total distance (cm) and average speed (cm/s) (Table 1). Female adult mice tended to show extreme values compared with male adult mice. In total, 8.8% and 15.2% of male and female mice, respectively, were hyperactive (determined by the total distance moved, >3500 cm). An average speed >7.0 cm/s was observed in 3.5% and 14.3% of male and female mice, respectively.
Discussion
In this study, female mice showed lower activity levels in terms of ambulation (including number of movements and movement time) and rearing (including number of vertical activities and vertical time) than male mice at 3 and 8 weeks of age. In addition, female adult mice showed shorter durations of ambulation and rearing than male adult mice. These effects were not observed in immature mice before weaning. Sex differences in exploratory behaviour of immature mice showed quantitative changes only. In adult mice, variables for rearing excluding rearing time showed quantitative changes only; however, variables for ambulation and rearing time showed both quantitative and qualitative (with different longitudinal pattern) changes. Since the results are based on data collected over 13 years, the genetic drift over generations might influence the results. Nevertheless, it seems that genetic drift had little influence on the results because the mice were purchased from the same major supplier for each study.
The differences reported in the present study might be influenced by/due to chronic psychosocial stressor (individual housing) exposure of the females. Numerous studies (Baer, 1971; Brain, 1975) have suggested that individually housed rodents, including mice, may exhibit physiological and behavioural changes when compared with their group-housed counterparts (ARRP, 2012). Nevertheless, several studies have reported no effects of individual housing on behavioural activities in mice (Arndt et al, 2009; Bartolomucci et al, 2009). It is also known that male and female offspring are differently affected when the dam is stressed prenatally (Archer & Blackman, 1971; Weinstock, 2007). In the present study, it seems that few effects of individual housing on exploratory behaviour were found between sexes since the number of defecations, indicative of emotionality, showed no differences between sexes in immature and adult mice.
Because the body size of mice is characterized by sexual dimorphism (Korkman, 1951; Eisen & Hanrahan, 1972), these changes in exploratory behaviour may be caused by differences in the body weight between the sexes. Wirth-Dzieciolowska et al. (2005) reported that selection for body weight induced differences in exploratory behaviour of mice. They found that the lighter group displayed a steady increase in behavioural activity during three sessions in an open-field test but displayed lower activity during the entire session than the other groups. Nevertheless, Tanaka (2010) reported that few variables associated with exploratory behaviour, except for rearing time in immature females, were influenced by the body weight of mice under natural conditions. The body weight of mice is not reported for the present study because the data are included in the previous study (Tanaka, 2010). Therefore, it seems that these changes in exploratory behaviour are caused by sex differences.
Valle (1970) reported that female rats were generally more active than male rats; however, the pattern of their activities did not differ from that of male rats in the open field. Masur (1972) found that male Wistar rats displayed more defecation and less ambulatory behaviour than female rats in the open field. Archer (1974) also reported that male Sprague–Dawley rats showed a lower latency time and more stationary sniffing than female rats. In mice, Archer (1977) found that males showed a lower latency time (C57BL and BALB/c) and more stationary sniffing (C57BL) than females. Archer (1975) suggested that male rats typically showed more defecation behaviour than female rats. However, in mice, neither sex typically shows more defecation behaviour than the other (Archer, 1975). In general, males display less ambulatory activity than females in both rats and mice. However, many studies have also reported no significant differences between the sexes.
Gray (1971) suggested that the uncertainty relating to sex differences in exploratory behaviour in mice may be caused by the predominant use of inbred strains in studies of these species. However, according to Beatty (1979), whether differences in the degree of inbreeding can explain this discrepancy remains unclear. Lightfoot (2008) reported female rodent daily activity levels that were 20% to more than 50% higher than those of males. Bronstein et al. (1975) observed that female Sprague–Dawley rats traversed a significantly greater distance during open-field testing than male rats, regardless of whether the animals had been handled or not. Gray (1979) suggested that similar to outbred rat strains, outbred mice strains exhibited the typical rat pattern of higher defecation rates and lower activity among males. Nevertheless, the results of the present study show the reverse in terms of ambulation and rearing in CD-1 (ICR) mice.
Valle & Bols (1976) found that male and female rats did not differ in their open-field behaviour at 30 days of age and that females showed more ambulation and rearing than males at 120 days of age. In addition, Beatty & Fessler (1976) reported that weanling male and female rats did not differ in terms of open-field activity; however, sex differences in all measures (ambulation, rearing and defecation) emerged by 50 days of age and persisted thereafter. Masur et al. (1980) reported that levels of defecation, ambulation and rearing were similar between male and female rats in the open field before 45 days of age. After 60 days of age, male rats showed more defecation and lower ambulation and rearing activity than female rats. Lynn & Brown (2009) found that male and female rats did not differ in terms of open-field activity as early (22–24 days of age) and middle (38–40 days of age) adolescents; however, sex differences emerged by late adolescence (52–54 days of age). Slob et al. (1986) observed that intact female rats were more active than their male counterparts in the open field at 77 days of age but not at an earlier age. Nevertheless, the sex difference in adulthood failed to occur in animals gonadectomised at 1 or 21 days of age. It therefore seems that sex differences in open-field activity occur in rats after puberty. In this study, sex differences were observed in exploratory behaviour of mice at 3 and 8 weeks of age (approximately 21 and 56 days of age, respectively). It therefore seems that sex differentiation in exploratory behaviour occurs in mice at an earlier age than that in rats.
Broida & Svare (1984) reported that intact females were more active than their male counterparts and that although gonadectomy in adulthood reduces behavioural levels in both the sexes, it does not eliminate sex differences in spontaneous behaviour in Rockland–Swiss albino mice. Guttman et al. (1975) found that motor activity in the barrier apparatus, as measured by jumping, reached a peak during oestrus and decreased between metoestrus and dioestrus in female mice. Palanza et al. (2001) reported that proestrus mice were less sensitive to the decrease in exploratory propensity induced by individual housing than oestrus and dioestrus mice. It seems, therefore, that gonadal hormones and the oestrous cycle influence activity levels in female mice.
In the present study, sex differences in variables related to ambulation in adult mice showed quantitative and qualitative changes. Female adult mice tended to show extremes of movement compared with male adult mice. Therefore, it is possible that oestrous females displayed such extreme activities, even though female mice were not examined for oestrous cycle at behavioural testing in the present study. Even though the ‘female’ group included animals that showed extremes in terms of movements, the group as a whole still showed lower activity in terms of exploratory behaviour than males. It seems, therefore, that in CD-1 mice, nonoestrus females probably display less exploratory behaviour than males. From the results of this study, it appears that female CD-1 mice display less exploratory behaviour than males and that these sex differences in exploratory behaviour occur from the prepubertal period.
References
-
Animal Research Review Panel (ARRP):Guidelines for the
Housing of Mice in Scientific Institutions (Guideline 22). Animal
Welfare Unit (Australia), 2012 Available at
http://www.animalethics.org.au/__data/assets/pdf_file/0004/249898
/Guideline-22-mouse-housing.pdf. Accessed at 22 January 2013.
-
Archer J: Sex differences in the emotional behavior of
three strains of laboratory rat. Anim. Learn. Behav. 1974,
2, 43–48.
-
Archer J: Rodent sex differences in emotional and related
behavior. Behav. Biol. 1975, 14, 451–479.
-
Archer J: Sex differences in the emotional behaviour of
laboratory mice. Brit. J. Psychol. 1977, 6, 125–131.
-
Archer JE & DE Blackman: Prenatal psychological stress
and offspring behavior in rats and mice. Devel. Psychobiol. 1971,
4, 193–248.
-
Arndt SS, MC Laarakker, HA van Lith, FJ van der Staay, E Gieling,
AR Salomons, J van’t Klooster & F Ohl: Individual housing of mice – Impact on behaviour and stress
responses. Physiol. Behav. 2009, 97, 385–393.
-
Baer H: Long-term isolation stress and its effects on drug
response in rodents. Lab. Anim. Sci. 1971, 21, 341–349.
-
Bartolomucci A, S Parmigiani, L Gioiosa, G Ceresini & P
Palanza: Effects of housing social context on emotional behaviour and
physiological responses in female mice. Scand. J. Lab. Anim. Sci.
2009, 36, 87–95.
-
Beatty WW & RG Fessler: Ontogeny of sex differences in
open-field behavior and sensitivity to electric shock in the rat.
Physiol. Behav. 1976, 16, 413-417.
-
Beatty WW: Gonadal hormones and sex differences in
nonreproductive behaviors in rodents: Organizational and
activational influences. Horm. Behav. 1979, 12, 112–163.
-
Brain P: What does individual housing mean to a mouse? Life
Sci. 1975, 16, 187–200.
-
Broida J & B Svare: Sex differences in the activity of
mice: Modulation by postnatal gonadal hormones. Horm. Behav. 1984,
18, 65–78.
-
Bronstein PM, FD Wolkoff & MJ Levine:
Sex-related differences in rats’ open-field activity. Behav.
Biol. 1975, 13, 133–138.
-
Eisen, EJ & JP Hanrahan: Selection for sexual
dimorphism in body weight of mice. Aust. J. Biol. Sci. 1972,
25, 1015-1024.
-
Fujikoshi Y, T Kan & Y Hijikata: Applied Longitudinal
Analysis. 2008, Ohmsha Ltd, Tokyo. (in Japanese)
-
Gray JA: Sex differences in emotional behaviour in mammals
including man: Endocrine bases. Acta Psychol. 1971, 35,
29–46.
-
Gray JA: Emotionality in male and female rodents: a reply
to Archer. Brit. J. Psychol. 1979, 70, 425–440.
-
Guttman R, I Lieblich & R Gross: Behavioral correlates
of estrous cycle stages in laboratory mice. Behav. Biol. 1975,
13, 127–132.
-
Korkman N: Selection with regard to the sex difference of
body weight in mice. Hereditas 1957, 43, 665-678.
-
Lightfoot JT: Sex hormones’ regulation of rodent
physical activity: A review. Int. J. Biol. Sci. 2008, 4,
126–132.
-
Lynn DA & GR Brown: The ontogeny of exploratory
behavior in male and female adolescent rats (Rattus norvegicus). Devel. Psychobiol. 2009, 51, 513–520.
-
Martin P & P Bateson: Measuring Behaviour: an
introductory guide. 1990, Tokai University Press, Tokyo, pp.
102-122.
-
Masur J: Sex differences in “emotionality” and
behavior of rats in the open field. Behav. Biol. 1972, 7,
749–754.
-
Masur J, MT Schutz & R Boerngen: Gender differences in
open-field behavior as a function of age. Devel. Psychobiol. 1980,
13, 107–110.
-
Matsumoto K, B Cai, S Nakamura & H Watanabe: A new
system for the measurement and analysis of motor activity in mice.
Folia Pharmacol. Jpn. 1990a, 96, 31–39 (in Japanese).
-
Matsumoto K, B Cai, K Sasaki & H Watanabe.Methamphetamine- and apomorphine-induced change in spontaneous
motor activity using a new system to detect and analyze motor
activity on mice. J. Pharmacol. Methods 1990b, 24,
111–119.
-
National Research Council: Guide for the Care and Use of
Laboratory Animals. 1996, National Academy Press, Washington D.C.
-
Palanza P, L Gioiosa & S Parmigiani. Social stressing
mice: Gender differences and effects of estrous cycle and social
dominance. Physiol. Behav. 2001, 73, 411–420.
-
The Science Council of Japan: Guidelines for Proper Conduct
of Animal Experiments. 2006, Tokyo: The Science Council of Japan.
Available at: http://
www.scj.go.jp/ja/info/kohyo/pdf/kohyo-20-k16-2e.pdf. Accessed 10
January 2007.
-
Slob AK, T Huizer & JJ van der Werff Ten Bosch.
Ontogeny of sex differences in open-field ambulation in the rat.
Physiol. Behav. 1986, 37, 313–315.
-
Tanaka T: Biological factors influencing exploratory
behavior in laboratory mice, Mus musculus. Mammal Study
2010, 35, 139–144.
-
Valle FP: Effects of strain, sex, and illumination on
open-field behavior of rats. Amer. J. Psychol. 1970, 83,
103–111.
-
Valle FP & RJ Bols: Age factors in sex differences in
open-field activity of rats. Anim. Learn. Behav. 1976, 4,
457–460.
-
Weinstock M: Gender differences in the effects of prenatal
stress on brain development and behaviour. Neurochem. Res. 2007,
32, 1730–1740.
- Wirth-Dzieciolowska E, A Lipska & M Wesierska: Selection for body weight induces differences in exploratory behavior and learning in mice. Acta Neurobiol. Exp. 2005. 65, 243-253.