
Anxiolytic effect of Mozart music over short and long photoperiods as part of environmental enrichment in captive Rattus norvegicus (Rodentia: Muridae)
by J.N. Cruz1, D.D. Lima2, D.D. Dal Magro3 & J.G.P. Cruz3*
1Department of Medicine, University of the Extreme South of Santa
Catarina, Criciúma - SC, Brazil
2Department of Pharmacy, University of Joinville Region,
Joinville - SC, Brazil
3Department of Natural Sciences, Regional University of
Blumenau, Blumenau - SC, Brazil
 Correspondence: J.G.P. Cruz
Correspondence: J.G.P. Cruz
Department of Natural Sciences, Reginal University of Blumenau, Rua
Antônio da Veiga, 140,
Blumenau, 89012-900, Santa Catarina, Brazil
Tel +55 047 3321 0272
Fax +55 047 3321 0233
E-mail jgcruz@furb.br
Summary
Music is known to be able to elicit emotional changes, including anxiolytic effects on humans and animals. Photoperiod has also been reported to play an important role in the modulation of anxiety. In the present study, we examined whether the effect of music on anxiety is influenced by day length, comparing, short day (SD; 8:16 h light/dark) and long day (LD; 16:8 h light/dark) with controls (CD; 12:12 h light/dark). After 8 weeks of photoperiod treatment, rats were randomly assigned to 2 groups: silence and music. In the music group, rats were exposed to music 24 h before behavioral tests to quantify anxiety level. Exposure to Mozart music reduced anxiety in rats in the CD group. These effects of music were abolished by LD. Independently of music, rats exposed to SD exhibited higher levels of anxiety-like behavior than rats exposed to CD, in elevated plus-maze and open-field tests. The present findings suggest that the anxiolytic effects of Mozart music are photoperiod-dependent.
Introduction
In the wild, animals are exposed to an ever-changing array of sensory stimuli. Captive environments for laboratory rodents, by contrast, appear generally more impoverished in terms of sensory stimuli. Lately, some attention has been directed towards exploring the impact of stimulation of one or more senses (e.g. auditory, vision, smell) as a method of environmental enrichment (Baumans 2005, Arakawa et al. 2011). The goals of environmental enrichment are non-specific and relatively open to debate, but it is generally agreed that enrichment strategies should encourage more species-typical patterns of behavior, increase the ability to cope with challenges, enhance behavioral repertoire, increase positive use of the environment and/or reduce or eliminate aberrant patterns of behavior, e.g. stereotypies (Wells & Irwin 2008, Wells 2009).
That music can have a soothing effect on human psychology and behaviors is well known (Koelsch & Siebel 2005, Koelsch 2010); whether it is soothing or stimulating or stressful depends on the type of music, and the circumstances (Cruz et al. 2010, Cruz et al. 2011). Recognition of the benefits associated with music for human well-being has prompted recent research into the value of auditory stimulation as a means of enriching the environment of captive animals. Classical music also appears to influence the behavior and/or physiology of some species of captive animals in a manner suggestive of enhanced well-being. In addition, other studies show that music can lead to positive behaviors (Wells et al. 2008) and physiological benefits (Núñez et al. 2002, Sutoo & Akiyama 2004, Nakamura et al. 2007). The value of auditory enrichment has been studied in a variety of species, including birds (Reed et al. 1993), cattle (Uetake et al. 1997), horses (Houpt et al. 2000), dogs (Wells et al. 2002), fish (Papoutsoglou et al. 2007) and primates (Videan et al. 2007, Wells et al. 2008). Many of these studies report changes in the behavior and/or physiology of animals exposed to music recordings, radio broadcasts or ecologically relevant sounds; but, to our knowledge, there is no literature about whether this effect is influenced by photoperiod.
Animal models of depression and anxiety may provide a means to elucidate the effect of photoperiod on anxiety and depressive-like behavior in humans. However, there has been little research on the effect of season on affective behavior in animals. Prolonged exposure to a long photoperiod (14/10 h light/dark cycle) in rats was found to have an antidepressant effect, as measured by reduced immobility in a forced swim test comparable to treatment with antidepressants (Molina-Hernandez & Tellez-Alcantara 2000). Another study in an animal exposed to varying photoperiods in its natural environment, the Siberian hamster, showed that the males exposed to short days had behavior suggesting greater levels of anxiety and depression compared to those exposed to long days; this effect did not appear among the females (Prendergast & Nelson 2005). In contrast, exposure to short days decreased neophobic reactions in C3H/He and BALB/c mice, while long days amplified the neophobia only in the C3H/He mice relative to controls (Kopp et al. 1997).
Studies in rodents have indicated that there are gender differences in emotional behavior caused by environmental conditions. Rats exposed to short days exhibited higher levels of anxious-like behavior than rats exposed to long days, in an elevated plus-maze and open-field test (Benabid et al. 2008). Based on these findings, we hypothesized that enriching the auditory environment with music might influence the effects of photoperiod on anxiety.
Most of the procedures used to assess anxiety in rodents can also be used to assess the anxiolytic effects of various agents. Typically in these studies, animals are exposed to anxiogenic conditions, e.g. a novel environment (elevated plus-maze and open-field tests), in order to study the anxiolytic properties of compounds, such as benzodiazepine (Boufleur et al. 2012). The aim of our experiments was to test the hypothesis that exposure to Mozart’s music throughout short or long daylight periods would influence anxiety state. A battery of two different anxiety tests was used. These consisted of the elevated plus-maze and open-field tests.
Materials and methods
Subjects
A total of 60 male genetically heterogeneous albino Wistar rats (Rattus norvegicus), aged 3 to 5 months and weighing 220 to 310 g, were obtained from
the animal house of the Regional University of Blumenau. After arrival
in the vivarium of the laboratory, these animals were housed in groups
of five per opaque plastic cage (50 x 30 x 15 cm) with wood shaving
bedding and wire mesh tops. They were housed under a standard (12/12 h
light/dark; cycle lights on at 7:00 AM), in a temperature-controlled
environment (23 ± 1ºC), with a 50 dB background sound level, and
55±10% relative humidity. During the light and dark phases, the rats
were exposed to light intensities of approximately 325 and 0,025 lux,
respectively. During the entire experimental period, the animals
received commercial chow for rodents (Nuvital, Paraná, Brazil) and
filtered tap water ad libitum. The animals were acclimated to
the animal housing facilities for at least 1 week before the
experiments began. The experiments were performed in compliance with
the recommendations of the Brazilian Society of Neuroscience and
Behavior (SBNeC), which are based on the United States National
Institutes of Health
Guide for Care and Use of Laboratory Animals.
Experimental protocols
The rats were divided into three groups: group control (CD – 12:12 h
light/dark, lights on from 07:00 to 19:00 h), short day (SD – 8:16 h
light/dark, lights on from 07:00 to 15:00 h) and long day (LD – 16:8 h
light/dark, lights on from 07:00 to 23:00 h). During the light and
dark phases, the rats were exposed to a light intensity of
approximately 325 and 0.025 lux, respectively. These lux values were
chosen because they were the closest values (in our laboratory)
possible to natural daytime and nighttime light. All rats were exposed
to their respective photoperiod conditions for 8 weeks before testing.
In the behavioral tests, rats were randomly assigned to two groups:
silence and music, with n = 10 per group for each photoperiod
condition. The rats in the auditory enriched conditions were exposed
to music 24 h before behavioral tests, throughout both dark and light
cycles, and also during the tests. The music (i.e., Mozart’s
piano sonata, KV361, Largo, 8:35 min duration) was played repeatedly
on a compact disc player (Cruz et al., 2010; Cruz et al., 2011). The speaker had a frequency range of 100-16000 Hz. The silent room
was exactly the same as the room in which music was played but no
sound was made, with the exception of ambient noises such as those
produced by the air conditioner. The sound levels for the silence and
music groups were 50 dB (ambient noise) and 65-75 dB, respectively, in
the home cages and behavioral apparatus.
Behavioral tests
The animals were individually subjected to the elevated plus-maze
and open-field tests. All of the behavioral
procedures were conducted during the light phase (between 7:30 and
11:30 AM), in a sound-isolated room, when the rodents were less
active. To minimize possible circadian influences, the experimental
and control observations were alternated. The observer stayed in the
same room, approximately 1 m away from the apparatus.
Elevated plus-maze test
The apparatus consisted of two open arms (50 x 10 cm) and two enclosed
arms (50 x 10 x 40 cm) arranged such that the two arms of each type
were opposite to each other, with a central platform (10 x 10 cm). The
height of the maze was 50 cm. The rats were individually placed on the
central platform of the elevated plus-maze facing an open arm. During
a 5 min test period the following parameters were recorded by an
observer: time spent in the open arms, number of entries into the open
arms, time spent in the enclosed arms, number of entries into the
enclosed arms.
Open-field test
The open-field consisted of a black circular box (60 cm diameter, 50
cm height). Each rat was placed in the central area and allowed to
freely explore the apparatus for 5 min. In the open-field test, we
recorded total time spent in locomotion (i.e. movement from one
location to another), the time spent immobile (i.e. completely
immobile), the time spent rearing (i.e. the rat stood on its hind paws
with its body at greater than a 45° angle to the floor) and the time
spent grooming (i.e. repetitive movements of the front paws or mouth
on the fur).
Statistical analysis
All of the data are expressed as ± S.E.M. Each value reflects the mean
of 10 animals per group. The means were compared using analysis of
variance (ANOVA), followed by the Newman-Keuls multiple comparison
test. A probability level of 0.05 was used as the criterion for
statistical significance.
Ethical considerations
All experiments were approved by the local ethical committee of the
Regional University of Blumenau.
Results
Elevated plus-maze test
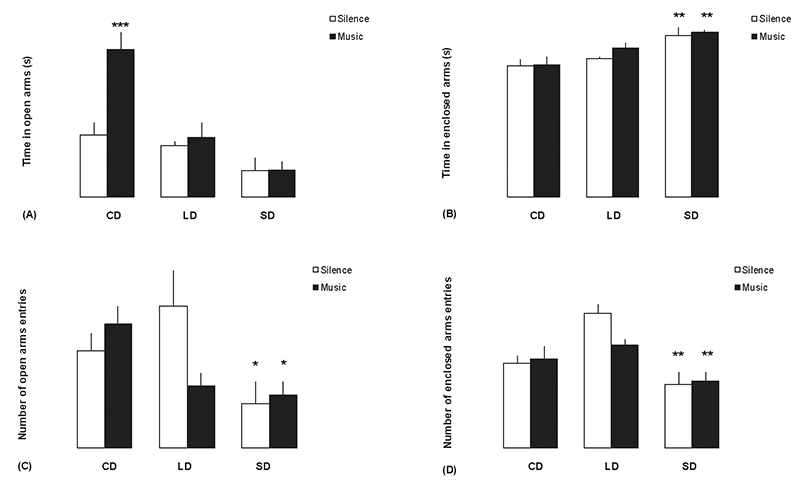
The ANOVA revealed significant differences between groups in the time
spent in open arms of the elevated plus-maze (F1,59 = 12.903; p<
0.001). In the control group (CD) music-exposed rats spent more time
in the open arms than rats exposed to silence (Figure 1A; p<
0.001). The photoperiod affected significantly the time in the
enclosed arms (F1,59 = 5.669; p< 0.001), and the number of open
(F1,59 = 3.345; p< 0.01) and enclosed (F1,59 = 8.106; p<
0.001) arms entries. SD rats exposed to music or silence spent more
time in enclosed arms (Figure 1B; p< 0.01), and entered less often
the open (Figure 1C; p< 0.05) and enclosed (Figure 1D; p< 0.01)
arms compared with rats in group CD/silence.
Open-field test
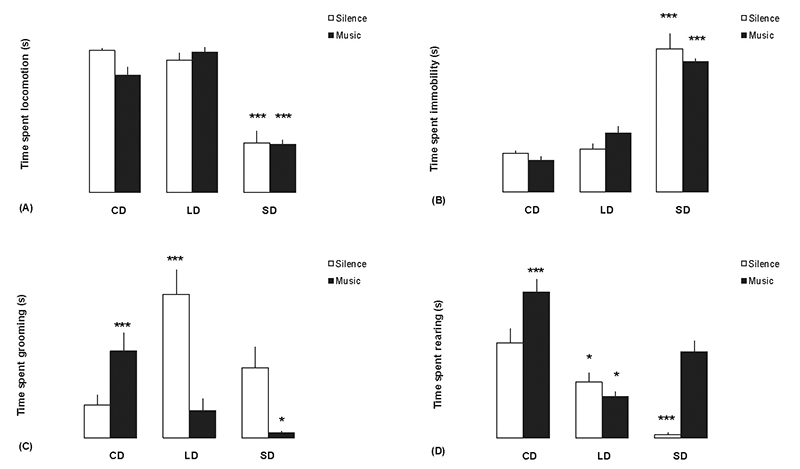
There were statistically significant differences between groups in the
total amount of time spent in locomotion (F1,59 = 38.228; p<
0.001) and immobility (F1,59 = 45.024; p< 0.001) in the
open-field. SD rats spent less time in locomotion (Figure 2A; p<
0.001) and more time immobile compared with group CD/silence (Figure
2B; p< 0.001). Photoperiod and music affected significantly the
time spent grooming (F1,59 = 9.349; p< 0.001; Figure 2C). The time
spent grooming in the presence of music increased in CD (p< 0,001)
and decreased in SD rats (p< 0.05) compared with group CD/silence.
Comparing the results for each silence group, LD rats spent more time
in the grooming (p< 0.001). The time spent rearing differed between
photoperiods (F1,59 = 23.774; p< 0.001; Figure 2D). Music-exposed
rats showed increased time spent rearing in the CD group (p< 0.001)
and decreased time in the LD group (p< 0.05) compared with group
CD/silence. However, Newman-Keuls test showed decreased time spent
rearing byrats exposed to silence in LD (p< 0.05) and SD groups
(p< 0.001) compared with the CD/silence rats.
|
|
Figure 1. Anxiety levels of rats in each sound condition (silence or music) under different photoperiods in the elevated plus-maze test. The rats were exposed to music 24 h before behavioral test, and also during the test. CD = Controls (12:12 h light/dark), LD = Long Day (16:8 h light/dark), and SD = Short Day (8:16 h light/dark). (A) Time in open arms. (B) Time in enclosed arms. (C) Number of open arms entries. (D) Number of enclosed arms entries. Vertical lines expressed as means ± SEM. Bars represent the mean of groups of 10 rats. Significant differences between intervention and CD/silence groups were observed:*p< 0.05; **p< 0.01; ***p< 0.001; ANOVA followed by Newman-Keuls test.
Click image to enlarge |
|
|
Figure 2. Differences in behaviors that may be indicative of anxiety levels in rats in each sound condition (silence or music) under different photoperiods in the open-field test. The rats were exposed to music 24 h before behavioral test, and also during the test. CD = Controls (12:12 h light/dark), LD = Long Day (16:8 h light/dark), and SD = Short Day (8:16 h light/dark). (A) Time spent in locomotion. (B) Time spent immobile. (C) Time spent grooming. (D) Time spent rearing. Vertical lines expressed as means ± SEM. Bars represent the mean of groups of 10 rats. Significant differences between intervention and CD/silence groups were observed:*p< 0.05; ***p< 0.001; ANOVA followed by Newman-Keuls test.
Click image to enlarge |
Discussion
The results of the study suggest exposure to music reduced anxiety in rats under the control (CD) photoperiod. The CD rats exposed to music spent more time in the open arms of the elevated plus-maze. The elevated plus-maze is a widely used animal model of anxiety that is based on two conflicting tendencies: the rodent’s drive to explore a novel environment and it’s aversion to open spaces. Thus anxious animals will spend most time in the closed arms while less anxious animals will explore open areas longer (Pellow et al. 1985). These effects of music were not seen in rats kept under short (SD) and long (LD) photoperiods. SD rats exhibited higher levels of anxious-like behavior than those in the LD and CD groups. The SD rats spent more time in enclosed arms and showed fewer entries into the enclosed or open arms of the elevated plus-maze, compared with the CD group/silence.
A number of studies have been performed on laboratory rodents to investigate the effects of light conditions on behavior, and similar results have been obtained. Indeed, rats and hamsters exposed to SD expressed greater levels of behaviors thought to be indicative of anxiety compared to those exposed to CD (Prendergast & Nelson 2005, Benabid et al. 2008). Together, these results indicate that changes in photoperiod length are associated with different levels of anxious-like behaviors. In terms of the relationship between the state of anxiety and inhibition of the natural tendency to explore novel and aversive environments, the data suggest that short photoperiod has a facilitator effect on anxious-like behaviors. In the open-field test, music increased exploratory activity only in the CD rats compared with other groups. Music-exposed CD rats showed increased time spent rearing and grooming. However, LD rats showed less time rearing (under music or silence conditions) but more time grooming when in the presence of silence. Rearing is often used to evaluate exploratory activity (Maier et al. 1988) and a decrease in exploratory drive has been observed in rats subjected to stressful events (Guimarães et al. 1993). Many studies suggest that grooming behavior is increased by a mild aversive stimulus; however, the response varies with the degree of fear and the habituation to the stressful situation (File et al. 1988). There was no significant difference between the two acoustic conditions in the CD group in time spent in locomotion in the open-field and in the number of open and closed arms entries in the elevated plus-maze. These results suggested that the observed effect of music was not due to changes in arousal level and basic locomotion.
Adaptation to SD also resulted in decreased locomotion and increased immobility in the open-field test. These results indicate that differences in photoperiod length were associated with different levels of anxious-like behaviors. Moreover, the transfer of rats to SD induces an expansion in the duration of nocturnal melatonin secretion and exposure to light in the dark phase suppresses melatonin secretion (Illnerova et al. 1978, 1983). A light pulse encountered at the midpoint of subjective night shortens the duration of melatonin production in Djungarian hamsters maintained in SD to the value observed in animals maintained in LD; the light pulse at midnight serves apparently as “dawn” (Hoffmann et al. 1981). The rhythmicity of the central nervous system which controls the rhythmic melatonin signal is also affected by photoperiod (Sumova et al. 1995), and disruption of biological rhythms by phase changes in central nervous system functioning can induce stress (Stewart et al. 1990). Sound is thought to be one of the environmental factors that affects stress, and music can also be a significant element for an enriched environment in comparison to silence. However, in the present study, music did not block anxious-like behaviors in SD rats. Independently of treatment (silence or music), rats exposed to SD exhibited higher levels of anxious-like behavior than rats adapted to CD, in the elevated plus-maze and in open-field tests. Total activity levels were also affected by exposure to SD.
Anxiety is a complex behavioral phenomenon resulting from dynamic interactions between an organism and its surrounding environment. An important and difficult question still pertains to the neural mechanisms that can account for the beneficial effect of Mozart music combined with CD on anxiety in rats. A variety of evidence suggests that the effects of light on the mammalian circadian system of rodents are mediated by glutamatergic mechanisms and that the N-methyl-D-aspartate (NMDA) receptor plays an important role in this regulation (Colwell 2001). A possible mechanism mediating these anxiolytic effects of Mozart music could involve modulation of the NMDA receptor (Cruz et al. 2011). Recently, much attention has been devoted to the glutamatergic system and to NMDA receptor antagonists in particular (Riazabermudo-Soriano et al. 2012). To our knowledge the current study is the first to demonstrate Mozart music effects under different day lengths and could suggest an additional mechanism of action underlying observations of music-induced anxiolytic actions. This may suggest that our efforts should be shifted to the pursuit of new alternative avenues, important for continuing to define distinguishable preclinical and clinical behavioral effects, for different types of anxiolytic therapy.
In summary, exposure to music reduced behaviors thought to be indicative of anxiety levels in rats in the CD group. This suggests that some types of music may enrich some environments in comparison to silence. However, music did not block behaviors thought to be indicative of anxiety in rats in the SD group. Further investigations are needed to explore how photoperiod affects response to music.
References
-
Arakawa H, S Cruz & T Deak: From models to mechanisms:
odorant communication as a key determinant of social behavior in
rodents during illness-associated states. Neuroscience and
Biobehavioral Reviews, 2011, 35 (9), 1916-1928.
-
Baumans V: Environmental enrichment for laboratory rodents
and rabbits: requirements of rodents, rabbits, and research. ILARJ
Journal / National Research Council, Institute of Laboratory
Animal Resources, 2005, 46 (2),
162-170.
-
Benabid N, A Mesfioui & A Ouichou: Effects of
photoperiod regimen on emotional behaviour in two tests for
anxiolytic activity in Wistar rat. Brain Research Bulletin, 2008,
75 (1), 53-59.
- Boufleur N, CT Antoniazzi, CS Pase, DM Benvegnú, RC Barcelos, GS Dolci, VT Dias, K Roversi, G Koakoskia, JG Rosa, LJ
-
Barcellos & ME burger: Neonatal tactile stimulation
changes anxiety-like behavior and improves responsiveness of rats to
diazepam. Brain Research, 2012, 1474 (1), 50-59.
-
Colwell CS: NMDA-evoked calcium transients and currents in
the suprachiasmatic nucleus: gating by the circadian system.
European Journal of Neuroscience, 2001, 13 (7),
1420-1428.
-
Cruz JG, DD Dal Magro & JN Cruz: Effects of classic
music as part of environmental enrichment in captive
Mus musculus (Rodentia: Muridae). Biotemas, 2010,
23 (2), 191-197.
-
Cruz JN, DD Lima, DD Dal Magro & JG Cruz: The power of
classic music to reduce anxiety in rats treated with simvastatin.
Basic and Clinical Neurocience, 2011, 2 (4), 5-11.
-
File SE, PS Mabbutt & JH Walker: Comparison of adaptive
responses in familiar and novel environments: modulatory factors.
Annals of the New York Academy of Sciences, 1988, 525 (1),
69-79.
-
Guimarães FS, EA Del Bel, CM Padovan, SM Netto & RT
Almeida: Hippocampal 5-HT receptors and consolidation of stressful
memories.Behavioural Brain Research, 1993, 58 (1-2),
133-139.
-
Illnerova H, M Backstrom, J Saaf, L Wetterberg & B Vangbo: Melatonin in rat pineal and serum; rapid parallel decline after
light exposure at night. Neuroscience Letters, 1978,
9 (2-3), 189-193.
-
Illnerova H, J Vaneek & K Hoffmann: Regulation of the
pineal melatonin concentration in the rat (Rattus norvegicus) and the Djungarian hamster (Phodopus sungorus).
Comparative Biochemistry and Physiology. A, Comparative Physiology,
1983, 74 (1), 155-159.
-
Hoffmann K, H Illnerova & J Vanecek: Effect of
photoperiod and of one minute light at night-time on the pineal
rhythm of N-acetyltransferase activity in the Djungarian
hamster (Phodopus sungorus). Biology of Reproduction, 1981,
24 (3), 551-556.
-
Houpt K, M Marrow & M Seeliger: A preliminary study of
the effect of music on equine behavior. Journal of Equine Veterinary
Science, 2000, 20 (11), 691-737.
-
Koelsch S: Towards a neural basis of music-evoked emotions.
Trends in Cognitive Sciences, 2010, 14 (3), 131-137.
-
Koelsch S & WA Siebel: Towards a neural basis of music
perception. Trends in Cognitive Sciences, 2005, 9 (12),
578-584.
-
Kopp C, R Misslin, E Vogel, MC Rettori, P Delagrange & B
Guardiola-Lemaître: Effects of day-length variations on emotional responses towards
unfamiliarity in Swiss mice. Behavioural Processes, 1997,
41 (2), 151-157.
-
Maier SE, P Vandenhoff & DP Crowne: Multivariate
analysis of putative measures of activity, exploration,
emotionality, and spatial behavior in the hooded rat (Rattus norvegicus). Journal of Comparative Psychology, 1988, 102 (4),
378-387.
-
Molina-Hernandez M & P Tellez-Alcantara:
Long-photoperiod regimen may produce antidepressant actions in the
male rat. Progress in Neuro-psychopharmacology & Biological
Psychiatry, 2000, 24 (1), 105-116.
-
Nakamura T, M Tanida, A Niijima, H Hibino, J Shen & K
Nagai: Auditory stimulation affects renal sympathetic nerve activity and
blood pressure in rats. Neuroscience Letters, 2007, 416 (2), 107-112.
-
Núñez MJ, P Mañá, D Liñares, MP Riveiro, J Balboa, J
Suárez-Quintanilla, M Maracchi, MR Ménez, JM Lóopez & M
Freire-Garabal: Music, immunity, and cancer. Life Sciences, 2002,
71 (9), 1047-1057.
-
Papoutsoglou SE, N Karakatsouli, E Louzos, S Chadio, D
Kalogiannis, C Dalla, A Polissidis & Z
Papadoupoulou-Faifoti: Effect of Mozart’s music (Romanze-Andante of “Eine
Kleine Nacht Musik”, sol major, K525) stimulus on common carp
(Cyprinus carpio L.) physiology under different light
conditions. Aquacultural Engineering, 2007, 36 (1),
61-72.
-
Pellow S, P Chopin, SE File & M Briley: Validation of
open: closed arm entries in the elevated plus-maze as a measure of
anxiety in the rat. Journal of Neuroscience Methods, 1985,
14 (3), 525-529.
-
Prendergast BJ & RJ Nelson: Affective responses to
changes in day length in Siberian hamsters (Phodopus sungorus). Psychoneuroendocrinology, 2005, 30 (5), 438-452.
-
Reed HJ, LJ Wilkins, SD Austin & NG Gregory: The effect
of environmental enrichment during rearing on fear reactions and
depopulation trauma in adult caged hens. Applied Animal Behaviour
Science,1993, 36 (1), 39-46.
-
Riazabermudo-Soriano C, MM Perez-Rodriguez, C Vaquero-Lorenzo
& E Baca-Garcia: New perspectives in glutamate and anxiety. Pharmacology,
Biochemistry, and Behavior, 2012, 100 (4), 752-774.
-
Stewart KT, AM Rosenwasser, H Hauser, JR Volpicelli & NT
Adler: Circadian rhythmicity and behavioral depression: effects of
stress. Physiology and Behavior, 1990, 48
(1), 149-155.
-
Sumova A, Z Travnickova, R Peters, WJ Schwartz & H
Illnerova: The rat suprachiasmatic nucleus is a clock for all seasons.
Proceedings of the National Academy of Sciences of the United States
of America, 1995, 92 (17), 7754-7758.
-
Sutoo D & K Akiyama: Music improves neurotransmission:
demonstration based on the effect of music on blood pressure
regulation. Brain Research, 2004, 1016 (2), 255-262.
-
Uetake K, JF Hurnik & L Johnson: Effect of music on
voluntary approach of dairy cows to an automatic milking system.
Applied Animal Behaviour Science, 1997, 53 (3), 175-182.
-
Videan EN, J Fritz, S Howell & J Murphy: Effects of two
types and genre of music on social behaviour in captive chimpanzees
(Pan troglodytes). Journal of the American
Association for Laboratory Animal Science, 2007, 46 (1),
66-70.
-
Wells DL: Sensory stimulation as environmental enrichment
for captive animals: a review. Applied Animal Behaviour Science,
2009, 118 (1-2), 1-11.
-
Wells DL, L Ggraham & PG Hepper: The influence of
auditory stimulation on the behaviour of dogs housed in a rescue
shelter. Animal Welfare, 2002, 11 (4), 385-393.
-
Wells DL & RM Irwin: Auditory stimulation as enrichment
for zoohoused Asian elephants (Elephas maximus). Animal
Welfare, 2008, 17 (4), 335-340.
- Wells DL, CL Mcdonald & JE Ringland: Color preferences in gorillas (Gorilla gorilla gorilla) and chimpanzees (Pan troglodytes). Journal of Comparative Psychology, 2008, 122 (2), 213-219.