
Temporary negative results of serological monitoring of rats with enzootic Haemophilus infection
by Ron Boot1*, Lia van de Berg1, Hein van Lith2
1Diagnostic Laboratory for Infectious Diseases, National Institute
of Public Health and the Environment, P.O. Box 1, 3720 BA Bilthoven,
The Netherlands;
2Division of Animal Welfare & Laboratory Animal
Science, Department of Animals in Science and Society, Faculty of
Veterinary Medicine, Utrecht University, P.O. Box 80166, 3508 TD
Utrecht, The Netherlands
 Correspondence: Ron Boot
Correspondence: Ron Boot
Diagnostic Laboratory for Infectious Diseases, National Institute of
Public Health and the Environment, P.O. Box 1, 3720 BA Bilthoven, The
Netherlands
Tel +31 30 274 3432
Fax: +31 30 274 4448
E-mail r.boot@hotmail.com
Summary
Infection by V-factor requiring Pasteurellaceae (Haemophilus
spp) bacteria frequently occurs in Specified Pathogen Free (SPF)
colonies of rats. Colonies may be monitored by methods that either
detect the bacteria or detect specific antibodies by for instance the
enzyme-linked immunosorbent assay (ELISA) and immunoblot (IB).
Breeding females in enzootically infected colonies transfer antibodies
to their young. Maternal immunity prevents the development of
antibodies by the pups yielding a transition period of low antibody
activity in the pups between waning maternal immunity and
seroconversion.
We studied antibody activity to Haemophilus antigens in young
rats delivered by Haemophilus infected dams by ELISA and IB
using antigens of Haemophilus strains H21 (from rat) and H35
(from guinea pig).
Young showed high ELISA and IB maternal antibody activity until
weaning, a decline to a low level between 9 and 12 weeks of age, and
then seroconversion.
As a consequence seroprevalence varied with age and during the
transition period H21 ELISA yielded a false negative result in
Haemophilus infected rats.
The age-related variation in antibody activity in enzootically
infected rodents has an impact on the development of sampling
strategies for serological health monitoring and interpretation of its
results.
Introduction
The Federation of Laboratory Animal Science Associations (FELASA)
recommends monitoring rodent colonies for all Pasteurellaceae
bacteria (Nicklas et al., 2002). Pasteurellaceae comprise
growth factor independent species such as Pasteurella
pneumotropica and bacteria that need X- (hemin) and/or
V-factor (nicotinamide adenine dinucleotide [NAD]) for growth (Olsen et al., 2005). The latter bacteria are traditionally called
Haemophilus spp.
Most Pasteurellaceae from rodents belong to the Rodent cluster which
comprises biotype Heyl and Jawetz P.
pneumotropica and V-factor dependent strains (Olsen et al., 2005).
Haemophilus spp frequently occur in contemporary laboratory
rat and guinea pig colonies (Nicklas 1989;
Nicklas et al., 1993; Boot et al., 1994/5).
Haemophilus infected rats of various strains usually show
hyperaemic bronchial lymph nodes, peribronchiolar hyperplasia and
multifocal interstitial pneumonia (Nicklas 1989;
Nicklas et al., 1993; Boot et al., 1999).
Haemophilus strain H21 has been found to induce tracheal
hyporeactivity to carbachol in mice and guinea pigs used in asthma
research as did P. pneumotropica and other growth
factor independent species (Boot et al., 1999).
Haemophilus spp may significantly reduce antibody development
to Streptococcus pneumoniae in rats presumably by
interference with the recruitment of neutrophil leucocytes (Craig et al., 2009). Importantly Haemophilus infection should be absent from
rats used for potency testing of Haemophilus influenzae type
b vaccine.
Infection by Pasteurellaceae including Haemophilus species
can be detected by culture, polymerase chain reaction (PCR) (Bootz et al., 1998; Boot et al., 2009) and by serology (Boot et al., 1994/5; Boot et al., 2005). We use host species specific antigen
panels for large scale serological testing of animal colonies and
confirm serological observations by culture and/or PCR. Based on
cross-reactivity studies (Boot et al., 1996/7;
Boot et al., 1999) the antigen panel for rats includes two
Haemophilus strains, namely rat strain H21 that is in the
Rodent cluster and guinea pig strain H35 that belongs to the
H. parainfluenzae-complex (Boot & Reubsaet, 2009).
In a study using young rats from a Haemophilus infected
breeding colony we observed a decline in antibody activity against
both Haemophilus antigens. This observation suggested waning
of maternal antibodies which prevent production of antibodies by the
young. Young rats will only seroconvert after maternal antibodies have
fallen below a threshold (Ziang & Ertl, 1992;
Sigrist, 2003). This implies a transition period with low
antibody activity in young rats which might have consequences for the
outcome of serological monitoring.
We studied antibody activity to Haemophilus antigens in young
rats born from Haemophilus infected dams by enzyme-linked
immunosorbent assay (ELISA) and immunoblot (IB). Young showed high
maternal antibody activity until weaning, a decline to a low level
between 9 and 12 weeks of age, and then seroconversion. The major
consequence of the age-related variation in antibody activity was a
low seroprevalence during the transition period and young were
temporarily negative by Haemophilus H21 ELISA.
Materials and methods
Ethical note
Animal experiments were approved by the institute’s Ethical
Committee on Animal experiments and were conducted in compliance with
national legislation which is based on European Community Directive
86/609/EEC.
Animals
Eight mated female HsdCpb:WU rats aged 6-7 months were obtained from a
Haemophilus infected breeding colony. They delivered 84 young
within a period of one week. Litter size ranged from 5-13. To study
antibody activity in individual young rats over time, four rats per
litter comprising 2 males and 2 females from 7 litters and 3 males and
1 female from 1 litter were sampled.
Housing
Mothers delivered in a type III macrolon® cage and raised their young
until weaning at 3 wks. At weaning four male and four female groups
were formed from male (n=17) and female (n=15) young rats from litters
1 and 2; 3 and 4; 5 and 6; and 7 and 8 respectively and housed in open
type III cages within the same animal room. The groups contained 3
(one group), 4 (six groups) or 5 (one group) rats. No other laboratory
animal species or rat strains were present in the animal room.
Samples
Dams were blood sampled at delivery and at weaning of their young (16
samples).
From each of the 32 young 8 consecutive blood samples were obtained at
3 week intervals from 3 to 24 weeks (256 samples). The remaining 52
littermates were blood sampled when sacrificed at birth (n=8), or at 1
(n=7), 2 (n= 7) or 3 weeks (n= 30) of age.
Samples from rats at birth and at 1 week of age were obtained after
decapitation.
All other samples were obtained under KRA [Ketamine (Alfasan, Woerden,
The Netherlands) 90 mg/kg intraperitoneally, Rompun (Bayer AG,
Leverkusen, Germany) 10 mg kg intraperitoneally, atropine (Vetinex
Animal Health, Bladel, The Netherlands) 0.05 mg/kg intraperitoneally]
anesthesia through puncture of the orbital plexus or cardiac puncture
when rats were sacrificed. Sera were stored at -20 °C until
testing.
Pharyngeal samples for culture and PCR analysis were obtained from the
dams at weaning of their young and from young sacrificed at 3 (n= 30)
and 24 weeks of age (n= 32).
Serology
ELISA
The ELISA was carried out as described previously using whole cell
antigens of Haemophilus strains H21 and H35 and various other
bacteria (Boot et al., 1995). ELISA activity was measured as
optical density (OD) minus the OD of the negative control. We included
serology using antigen of Bordetella bronchiseptica as a
representative of absent infections (negative control).
For the preparation of whole cell antigens both
Haemophilus strains and B. bronchiseptica were grown
on chocolate agar and plain sheep blood agar respectively for 18 hrs
at 37 °C under 7.5% CO2, harvested in 0.07 M phosphate buffered saline
(PBS) pH 7.2, washed 3 times by centrifugation (300 g, 20 min) in PBS
and finally resuspended in PBS containing 0.01% (w/v) merthiolate. The
antigens were adjusted to 7.5 g/l of protein and coated to
polyvinylchloride flat bottomed microtitre plates (Titertek type III,
Flow Lab. Inc., Mc Lean, VA, USA).
Sera were tested as 1:50 dilutions for IgG antibodies using
horseradish peroxidase conjugated sheep anti-rat IgG (Sigma Aldrich
Chemie BV, Veenendaal, The Netherlands) as the secondary antibody.
Autologous positive control sera raised by immunisation and negative
control sera (from Pasteurellaceae- and
B. bronchiseptica-free rats) were run in each test.
Immunoblot (IB)
In 6 litters young showed a clear age-related variation in ELISA
antibody activity.
For these 6 dams serum collected at delivery and serum from one young
per litter at 3 weeks (‘peak’), 12 weeks
(‘dip’) and 24 weeks (‘peak’) were tested for
antibodies to both Haemophilus antigens by IB.
The IB was essentially carried out as described for
Streptobacillus moniliformis (Boot et al., 2006).
Haemophilus cells were grown, harvested and washed as
described above and were boiled (150 mg/l protein) in tap water for 3
min. Proteins were electrophorized on 10% polyacrylamide gels in
sodium dodecyl sulfate (SDS). Antigens were electrophoretically
transferred onto nitrocellulose membrane (Protran BA83, Schleicher
& Schuell, Dassel, Germany). Blots were incubated overnight at 4
°C in 0.05% Tween20/phosphate buffered saline (PBS/Tween) containing
1% low fat milk (Protifar, Nutricia, Zoetermeer, The Netherlands) [and
frozen at -20 °C until use]. Immunodetection was performed by
incubation of the blots with serum samples diluted 1:50 in PBS/Tween
for 60 min. at room temperature. After washing in PBS/Tween, the blot
was incubated with peroxidase conjugated sheep anti-rat IgG (Sigma
Aldrich Chemie BV, Veenendaal, The Netherlands) 1:2,000 diluted in
PBS/Tween. After washing in PBS/Tween, tetramethyl benzidine/dioctyl
sodiumsulfosuccinate (TMB/ DONS 0.06% and 0.2% respectively in
dimethyl sulfoxide) was added as substrate. Positive and negative
control sera and molecular weight marker with a range 10 to 250 kDa
(Kaleidoscope Prestained Standard 161-0375, Biorad Laboratories Ltd,
Veenendaal, The Netherlands) were included in each run.
Culture
Pharyngeal samples were cultured for respiratory bacterial infections (notably by Pasteurellaceae and B. bronchiseptica) on plain sheep blood agar and chocolate agar which were incubated for 24-48 hours at 37°C and read. Haemophilus suspected growth was identified and typed by the API NH system.
PCR analysis
Pharyngeal samples were PCR tested for Pasteurellaceae infection using the primer set as described by Bootz et al. (Bootz et al., 1998; Boot et al., 2009).
Statistical analysis
Optical density (OD) measured by ELISA with each of the 3 antigens was
expressed as a percentage of the activity of the autologous positive
control serum [(OD Sample / OD C+) x 100%) or in short: S/C+ %]. In
this way day to day variations in ELISA outcome due to time and
temperature differences in successive runs were minimized.
For the calculation of seroprevalence(s) S/C+ percentages
> 30 were considered positive. Although this percentage may
seem an arbitrary cut-off level, our ELISA results generally agree
with the outcome of culture and/or PCR for colonies of infected and
uninfected rats of various strains with different background (enteric)
flora.
The experimental (and statistical) unit is the entity that can be
assigned at random to one of the treatments, independently of all
other experimental units. Any two experimental units must be able to
be assigned to different treatments (Festing & Altman, 2005). This implies that in this study the dam is the experimental unit.
All statistical analyses were carried out according to Petrie and
Watson (Petrie & Watson, 1995) using a SPSS
® for Windows (version 15.0) computer program (SPSS Inc., IL, USA). Two-sided, exact (i.e. for the non-parametric
tests) (Mundry & Fischer, 1998) probabilities were
estimated throughout. The probability of a Type I error < 0.05 was
taken as the criterion of significance. The antibody activity data
were continuous and were summarized as means with standard deviation
(SD). The Kolmogorov-Smirnov one sample test was used to check
Gaussianity of the data. All results were normally distributed. In
addition, percentages of positive test results (seroprevalence) in the
colony over time were calculated. To assess the relationship between
maternal antibody activity at delivery or after 3 weeks and in
offspring at 0 or 3 weeks of age Pearson’s linear correlation
coefficients (r) were calculated; significance was assessed by a
two-tailed test based on the t statistic. Differences in
ELISA antibody activity between dams at delivery or after 3 weeks and
young at 0 or 3 weeks of age, were evaluated with the paired
Student’s t test. Antibody activity data were subjected
to an univariate repeated measures ANOVA with a Huynh-Feldt correction
and (depending on the comparison) between-subject factors comprising
antigen, gender and time. In the analysis type IV sums of squares were
used. Homoscedasticity was tested using the Levene’s test,
which is a powerful and robust test based on the F statistic (Lim & Lo, 1996).
IB reactivity to the Haemophilus antigens was for all
individual sera calculated using the formula [a.1] + [b.2]+ [c.3] in
which a, b, c is the number of immunoreactive antigens showing a
reaction scored as 1 (light); 2 (medium) or 3 (heavy). For both
Haemophilus antigens the Spearman’s rank correlation
coefficient (Rs) was calculated between IB
reactivity scores and ELISA reactivity; significance was assessed by a
two-tailed test based on the t statistic.
Results
Bacteriology
Haemophilus spp infection was detected in all dams, in 23 out
of 30 young at weaning and in 29 out of 32 young at the end of the
study by culture and in all animals by PCR.
B. bronchiseptica was not cultured from any animal.
Haemophilus strains were of a variety of API NH biotypes
(data not shown).
Serology
Transmission of maternal antibodies
At delivery (week 0) and after 3 weeks dams showed appreciable ELISA
antibody activity to both Haemophilus antigens (Table 1) and
most young had at 0, 2 and 3 weeks high antibody activity to the H21
antigen and all young had high activity to the H35 antigen. At
delivery and after 3 weeks the antibody activity to both
Haemophilus antigens was significantly higher in sera from
the mothers than from their young. Antibody activity to both antigens
in young and their respective mothers was significantly correlated
(Table 2).
Neither dams nor their young showed appreciable antibody activity to
B. bronchiseptica (Table 1).
Table 1. ELISA antibody activity to
Haemophilus strains H21 and H35, and
B. bronchiseptica antigens in sera from
Haemophilus infected rat dams and their young.1
| Dams (n = 8)2 at | Young (n = 8)2 at age of | ||||
| Antigen | delivery | after 3 weeks | 0 weeks | 2 weeks | 3 weeks |
| Haemophilus H21 | |||||
| Number (percentage) of positive experimental units | 6 (75.0) | 6 (75.0) | 5 (62.5) | 6 (85.7)3 | 6 (75.0) |
| Antibody activity (mean ± SD) | 59.4 ± 28.7 | 70.0 ± 35.4 | 40.4 ± 25.2 | 74.9 ± 25.6 | 46.5 ± 23.3 |
| Dams versus young (i.e. at week 0 of 3) | P = 0.000 | P = 0.004 | |||
| Haemophilus H35 | |||||
| Number (percentage) of positive experimental units | 8 (100) | 8 (100) | 8 (100.0) | 7 (100)3 | 8 (100.0) |
| Antibody activity (mean ± SD) | 117.0 ± 33.6 | 142.8 ± 15.8 | 98.0 ± 42.7 | 138.4 ± 26.7 | 102.7 ± 25.6 |
| Dams versus young (i.e. week 0 of 3) | P = 0.002 | P = 0.000 | |||
| B. bronchiseptica | |||||
| Number (percentage) of positive experimental units | 1 (12.5) | 1 (14.3)3 | 0 (0.0) | 1 (14.3)3 | 1 (12.5) |
| Antibody activity (mean ± SD) | 11.8 ± 12.1 | 12.4 ± 12.0 | 6.3 ± 8.1 | 16.1 ± 13.8 | 12.6 ± 13.5 |
| Dams versus young (i.e. week 0 of 3) | P = 0.057 | P = 0.589 | |||
1ELISA = enzyme-linked immunosorbent assay.
3Number of experimental units.
3Based on 7 experimental units.
3P values in paired Student’s t test. Significant
differences (P < 0.05) are indicated in bold characters. Note that
a P value of 0.000 does not mean that it is zero, only that
it is less than 0.0005.
Table 2. Associations between Haemophilus infected rat dams and their young with respect to ELISA antibody activity to Haemophilus strains H21 and H35, and B. bronchiseptica antigens in serum.1
| Antigen | Young at age of 0 weeks and dams at delivery | Young at age of 3 weeks dams after 3 weeks | |
| Haemophilus H21 | |||
| Pearson’s linear correlation coefficient | r = 0.956 | r = 0.933 | |
| P value | P = 0.000 | P = 0.001 | |
| Number of experimental units | n = 8 | n = 8 | |
| Haemophilus H35 | |||
| Pearson’s linear correlation coefficient | r = 0.984 | r = 0.823 | |
| P value | P = 0.000 | P = 0.012 | |
| Number of experimental units | n = 8 | n = 8 | |
| B. bronchiseptica | |||
| Pearson’s linear correlation coefficient | r = 0.843 | r = 0.975 | |
| P value | P = 0.009 | P = 0.000 | |
| Number of experimental units | n = 7 | n = 7 | |
1ELISA = enzyme-linked immunosorbent assay. All associations are significant (P < 0.05) and are indicated in bold characters. Note that a P value of 0.000 does not mean that it is zero, only that it is less than 0.0005.
All litters at birth showed appreciable ELISA antibody activity (i.e ≥
30% of the positive control serum) to the H35 (P = 0.003,
paired Student’s t test, n = 8), but not to the H21
antigen (P = 0.283, paired Student’s t test, n
= 8) (Fig. 1, panels a and b). ELISA antibody activity to
B. bronchiseptica in litters at birth was on average below
the threshold (P = 0.000, paired Student’s
t test, n = 8, Fig. 1, panel c).
All litters - except litter 3 for which no samples were available at
the age of 1 and 2 weeks - showed an increase in antibody activity to
Haemophilus antigens reaching a peak at 2 weeks and a
subsequent decline to a low activity plateau between 9 and 15 weeks.
From 15 weeks all litters showed an increase in antibody activity
(seroconversion) to Haemophilus antigens and the highest
activities were reached at end of the study (24 weeks). As a
consequence there was a significant time-effect for both
Haemophilus antigens in the repeated measurements ANOVA
(Table 3). Male and female young of a litter showed a similar pattern
of antibody activity which seemed unaffected by the formation of male
and female groups by mixing animals from different litters. ELISA
antibody activity to B. bronchiseptica was in the young
between weeks 0 and 24 on average below the threshold.
ELISA activity against the Haemophilus H35 antigen was in all
samples, and hence in all litters, higher than that to the
Haemophilus H21 antigen. The ratio of ELISA activity to the
H35 and the H21 antigens varied per litter and sampling time.
Antibody activity to B. bronchiseptica was at all sampling
times at a much lower level than the activity to both
Haemophilus antigens and only in one litter was a slighty
elevated antibody activity to B. bronchiseptica found at 2
weeks of age (Fig. 1c).
Seroprevalence
Average antibody levels and percentages of positive test results
(seroprevalence), for rats from all litters, to the two
Haemophilus antigens at each time point is shown in Fig. 2.
For both Haemophilus antigens there was an age related
variation in antibody levels and hence seroprevalence. The variation
was most striking for the Haemophilus H21 ELISA with a dip
between 6 and 15 weeks of age and a 0% seroprevalence at 9 weeks. The
seroprevalence measured by the Haemophilus H35 ELISA showed a
dip at 9-12 weeks but seroprevalence did not fall below 60%.
|
|
Figure 1. ELISA antibody activity to Haemophilus strains H21 and H35 and to B. bronchiseptica antigens in litters delivered by 8 Haemophilus naturally infected rat dams.
Click images to enlarge |
|
|
Figure 2. Variation in antibody activity to Haemophilus antigens in rats delivered by Haemophilus infected dams, leads to variation in seroprevalence
Click image to enlarge |
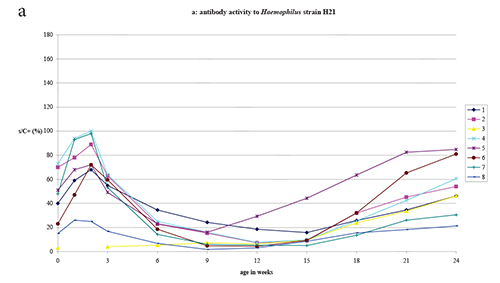
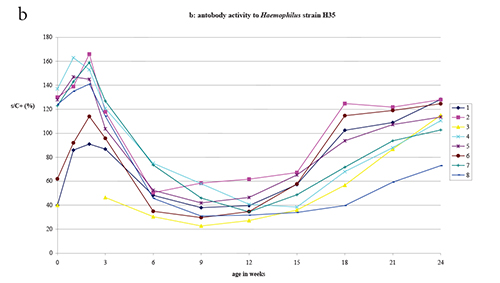
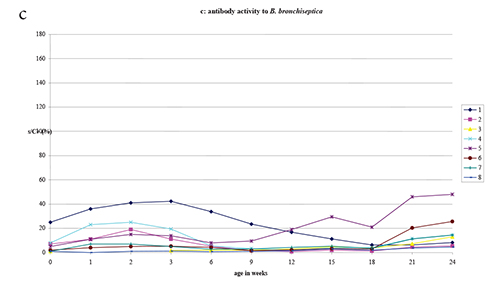
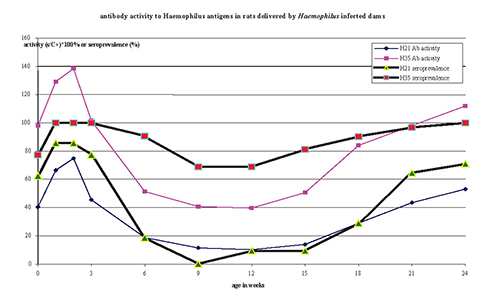
Table 3. P values in the different repeated measures ANOVA’s.1
| Antigen | Weeks | A2 | G | T | AxG | AxT | GxT | AxGxT |
| H21, H35, B. bronchiseptica | 3, 6, 9, 12, 15, 18, 21, and 24 | 0.000 | 0.472 | 0.000 | 0.671 | 0.000 | 0.592 | 0.092 |
| H21, H35 | 3, 6, 9, 12, 15, 18, 21, and 24 | 0.000 | 0.497 | 0.009 | 0.560 | 0.431 | 0.008 | 0.495 |
| H21 | 3, 6, 9, 12, 15, 18, 21, and 24 | - | 0.255 | 0.000 | - | - | 0.145 | - |
| H35 | 3, 6, 9, 12, 15, 18, 21, and 24 | - | 0.824 | 0.000 | - | - | 0.436 | - |
| B. bronchiseptica | 3, 6, 9, 12, 15, 18, 21, and 24 | - | 0.512 | 0.171 | - | - | 0.399 | - |
| H21, H35, B. bronchiseptica | 0, 1, 2, 3, 6, 9, 12, 15, 18, 21, and 24 | 0.000 | - | 0.000 | - | 0.001 | - | - |
| H21, H35 | 0, 1, 2, 3, 6, 9, 12, 15, 18, 21, and 24 | 0.000 | - | 0.000 | - | 0.123 | - | - |
| H21 | 0, 1, 2, 3, 6, 9, 12, 15, 18, 21, and 24 | - | - | 0.000 | - | - | - | - |
| H35 | 0, 1, 2, 3, 6, 9, 12, 15, 18, 21, and 24 | - | - | 0.001 | - | - | - | - |
| B. bronchiseptica | 0, 1, 2, 3, 6, 9, 12, 15, 18, 21, and 24 | - | - | 0.232 | - | - | - | - |
1Significant effects (P < 0.05) are indicated in bold
characters. Note Note that a P value of 0.000 does not mean that it is
zero, only that it is less than 0.0005.
2A = effect of antigen; G = effect of gender; T = effect of
time; AxG = interaction effect between antigen and gender; AxT =
interaction effect between antigen and time; GxT = interaction effect
between gender and time; AxGxT = interaction effect between antigen,
gender, and time.
Table 4. Immunoblot reactivity against antigens of Haemophilus strains H21 and H35 in naturally Haemophilus infected rat dams and their young.
| Young | ||||
| Haemophilus antigen | Dams at birth | 2 wks | 12 wks | 24 wks |
| strain H21 | 29* | 21,00 | 5 | 26 |
| strain H35 | 22 | 14,00 | 2 | 18 |
Immunoblot
As shown in Fig. 1a-b, 6 litters young showed a clear age-related
variation in ELISA antibody activity. Sera from the 6 corresponding
dams detected various immunodominant antigens in both
Haemophilus strains and banding patterns differed between
dams. The strain H21 antigens most frequently detected measured 10-15,
37, 44 and 100 kDa; the H35 antigens most frequently detected were of
10-15, 40, 57 and 100 kDa size. The banding pattern detected by sera
from dams and their young at 2 weeks of age appeared similar.
Total IB reactivity found in sera from young at 2 weeks of age (Table
4) diminished to a low level at 9-12 weeks (ELISA dip) but IB
reactivity reached maternal levels in sera taken from young rats at
the end of the study. IB reactivity scores for both
Haemophilus antigens were significantly correlated with ELISA
reactivity (Haemophilus H21: Rs = 0.717,
P = 0.000081; Haemophilus H35: Rs = 0.696, P = 0.000157).
Discussion
Transmission of maternal immunity in rat
The transfer of maternal antibodies from rat dams to offspring occurs
partly in utero but mainly postnatally via the mammary gland
in the first two weeks of life (Brambell, 1970;
Bainter, 2007). We found anti-Haemophilus antibody
activity immediately after birth (Fig. 1a-b) in offspring delivered by
Haemophilus infected dams and a rapid increase in antibody
activity in littermates in the first two weeks of life (Table 1 and
Fig. 1a-b). Antibody activity in young at 0 and 3 weeks of age
significantly correlated with levels measured in the dams at delivery
and 3 weeks later (Table 2); this agrees with findings in various
other species of animal (Grindstaff et al., 2003).
ELISA activity measured by the B. bronchiseptica antigen
represented absent infections in our study. Usually some background
ELISA activity is measured in uninfected animals due to
cross-reactivity with non-pathogenic bacteria entering the body e.g.
via the food. This aspecific activity (Fig. 1c) is also transmitted to
the offspring but it is at a much lower level than activity against
infecting bacteria such as Haemophilus spp (Fig. 1 a, b).
Diminishing maternal antibody in young rats and
seroconversion
From 2 weeks of age anti-Haemophilus antibody activity
declined to a plateau that was reached at about 6-9 weeks and from
12-15 weeks returned to higher levels (Fig. 1 a-b). A similar pattern
of decline and seroconversion was found in young delivered by
Corynebacterium kutscheri infected Wistar-Lewis,
Brown Norway (BN) and Fisher rats (Suzuki et al., 1988) and
by Mycoplasma pulmonis infected LEW rats (Cassell et al., 1986).
The M. pulmonis infected rats seroconverted within 2-4 months
whereas the C. kutscheri infected rats remained seronegative
for about 6 months. The long delay to measurable seroconversion to
C. kutscheri is likely due to the fact that antibodies to the
bacterium were measured by an agglutination assay (Suzuki et al., 1988) which is less sensitive than ELISA used to measure activity against
M. pulmonis or Haemophilus antigens (Cassell et al., 1988).
At the end of our study (24 weeks) young showed ELISA antibody
activities similar to those found in their dams, which is not
surprising as dams were 6-7 months of age when they delivered their
young.
Immunoblot
That IB reactivity (Table 4) followed variation in ELISA activity and
that activities measured by both assays were found to correlate was to
be expected. Pasteurellaceae including
Haemophilus spp are Gram negative bacteria with a complex
antigenic ‘make up’ comprising lipopolysaccharides and
proteins (Fenwick, 1995). Rodents infected by
P. pneumotropica raise ELISA antibodies against all these
kinds of antigen (Manning et al., 1989) of which those to
immunodominant proteins can be measured by IB.
Age-related IB reactivity has also been found in natural
Pneumocystis carinii infection in rats which showed
antibodies to four major antigens up to 4 weeks, but not at 8 weeks,
whereas later IB antibody activity returned as did ELISA activity (Hong et al, 1995). IB is used as a confirmatory test for ELISA results for instance
in monitoring infection by C. piliforme (Motzel et al., 1991), M. pulmonis (Simecka & Cassell, 1987) and
viral infections. Age of the animals should be considered in the
interpretation of IB test results.
By ELISA the serological behaviour of the young was similar within a
litter and apparently not affected by the formation of male and female
groups at weaning, whereas differences were seen between the litters
(Fig 1 a-b). Similar findings were reported for ELISA reactivity in
M. pulmonis naturally infected rat litters (Cassell et al., 1988). This supports observations that the antibody response to both
M. pulmonis (Simecka & Cassell, 1987) and
Haemophilus infection (Boot et al., 2005) is in rats
at least partly under genetic control.
Litter and strain related differences in IB reactivity complicate the
interpretation of IB test results.
Seroprevalence and consequences for health monitoring
An important implication of the age-related variation in ELISA
antibody levels is an age- related variation in seroprevalence
measured for the group (or colony) (Fig. 2).
The prevalence measured by a given assay determines the sample size
needed to detect at least one positive animal in a sample (Jonas,
1976). FELASA recommends a sample size of 10 animals (or samples) for
the periodic serological health monitoring of rodent colonies which
means that infections with a prevalence under 30% will not be detected
(Nicklas et al., 2002).
The seroprevalence measured by our Haemophilus H21 ELISA was
under 30% in rats aged 6 to 18 weeks and so the assay, if used alone
at these times, would have yielded a negative health monitoring record
for the colony. The seroprevalence measured by the H35 ELISA remained
above 60% (Fig. 2).
Selection of animals for health monitoring by serology
Our findings lead us to consider that the selection of animals might
be adapted to different situations.
To evaluate a colony of unknown infection history one should test
samples from different age categories: around weaning, 9-12 weeks and
> 24 weeks (6 months). Males and females seem to be equally
suitable for testing but it is advisable to sample animals from
different litters. An age-related antibody activity pattern as in Fig.
2 indicates enzootic infection. Antibody activity on a low level as
measured to B. bronchiseptica in rats < 6 weeks and >
15 weeks (Fig. 1c) suggests aspecific background activity. Antibody
activity at the age of 9-12 weeks clearly exceeding activity in other
age groups indicates recent infection. Positive and unequivocal
serological findings should be confirmed by culture and/or PCR. If
infection cannot be detected by other means the colony may be
considered uninfected.
Subsequent testing of such an uninfected rat colony might be limited
to animals aged 9-12 weeks as they will likely seroconvert upon recent
infection. In rats exposed to various different pathogenic bacteria in
a model mimicking natural infection seroconversion was found from
weaning (Boot, 2001).
Rederivation of a rat colony to (re)establish SPF status can involve
hysterectomy or embryo transfer. Hysterectomy derived rats foster
nursed to SPF dams should be tested (together with sera from donor
animals obtained at hysterectomy) at weaning, 9-12 weeks and for
instance at 15-18 weeks. Prenatally transmitted maternal antibodies
will decline both in infected and in uninfected young. An increase in
antibody activity from 15 weeks indicates active infection and hence
failure of the rederivation efforts. Difficulties in the
interpretation of serological findings can be circumvented by
rederiving colonies by embryo transfer to seronegative fosters.
It is obvious that quantification of antibody activity will be helpful
in the interpretation of serological findings. We expressed ELISA
antibody activity of our samples as a percentage of the activity of
the positive control sample. In assays such as IFA (indirect
fluorescent antibody) and agglutination titres should be determined
We showed that Haemophilus enzootically infected rats have an
age-related variation in antibody activity to the infection. Such a
variation leads to an age related variation in seroprevalence. This
phenomenon will also occur with other enzootic infections in rats.
Temporary low antibody activity due to the decline of maternal
antibodies is also to be expected in mice as they resemble rats in the
transmission of maternal immunoglobulins (Brambell, 1970).
We conclude that the likelihood of detection of infections by serology
in an enzootically infected colony depends on age of the animals
tested.
References
-
Bainter K. Transmission of antibodies from mother to young:
evolutionary strategies in a proteolytic environment. Vet. Immunol.
Immunopathol. 2007, 117, 153-161.
-
Boot R, H Thuis & MA Koedam. Infection by V-factor
dependent Pasteurellaceae (Haemophilus) in rats. J. Exp.
Anim. Sci. 1994/5, 37, 7-14.
-
Boot R, H Thuis, JL Veenema & RHG Bakker. An
enzyme-linked immonosorbent assay (ELISA) for monitoring rodent
colonies for Pasteurella pneumotropica antibodies. Lab.
Anim. 1995, 29, 307-313.
-
Boot R, H Thuis & JL Veenema. Serological relationship
of some V factor dependent Pasteurellaceae (Haemophilus sp)
from rats. J. Exp. Anim. Sci. 1996/7, 38, 147-152.
-
Boot R, HCW Thuis & JL Veenema. Serological
relationship of some V-factor dependent Pasteurellaceaea (Haemophilus
sp.) from guinea pigs and rabbits. Lab. Anim. 1999, 33:
91-94.
-
Boot R, J Garsen, MA Koedam & HCW Thuis.
Haemophilus sp infection is common in ‘pasteurella
free’ guinea pigs and must be known to interprete results of
pulmonary hypersensitivity studies. Revista de Ciencia 1999,
23, 21.
-
Boot R. Development and validation of ELISAs for monitoring
bacterial and parasitic infections in laboratory rodents and
rabbits. Scand. J. Lab. Anim. Sci. 2001, 28, 44-50.
-
Boot R, L van de Berg, H van Lith & JL Veenema. Rat
strains differ in antibody response to natural
Haemophilus spp infection. Lab. Anim. 2005, 39,
413-420.
-
Boot R, L van de Berg & MJ Vlemminx. Detection of
antibodies to Streptobacillus moniliformis in rats by an
immunoblot procedure. Lab. Anim. 2006, 40, 447-455.
-
Boot R, MJ Vlemminx & FAG Reubsaet. Comparison of
polymerase chain reaction primer sets for amplification of rodent
Pasteurellaceae. Lab. Anim. 2009, 43, 371-375.
-
Boot R & FAG Reubsaet. PCR is superior in detection of
Haemophilus infection in rats and guinea pigs. Scand. J.
Lab. Anim. Sci. (submitted feb. 2009).
-
Bootz F, S Kirschnek, W Nicklas, SK Wyss & FR Homberger.
Detection of Pasteurellaceae in rodents by polymerase chain reaction
analysis. Lab. Anim. Sci. 1998, 48, 542-546.
-
Brambell FWR. The transmission of passive immunity from
mother to young. Amsterdam, North-Holland Publishing Co. 1970.
-
Cassell GH, NF Cox &, JK Davis. State-of-the-art
detection methods for rodent Mycoplasmas. Pp 143-160 in:
Complications of Viruses
and Mycoplasmas in Rodents to Toxicology Research
and Testing (Hamm TE ed.) Hemisphere Publishing, Washington
DC 1986
-
Craig A, J Mai, S Cai & S Jeysaseelan. Neutrophil
recruitment of the lungs during bacterial pneumonia. Infect. Immun.
2009, 77, 568-575.
-
Davis J, GH Cassell, G Gambil, N Cox, H Watson & M
Davidson.
Diagnosis of murine mycoplasmal infections by enzyme-linked
immunosorbent assay. Isr. J. Med. Sci. 1987, 23, 717-722.
-
Fenwick B. Liposaccharides and capsules of the HAP group
bacteria. Pp. 75-87 in: Haemophilus, Actinobacillus and Pasteurella.
(Donachie W, Lainson FA, Hodgson JC eds.) Plenum Press. New
York-London 1995.
-
Festing MF & DG Altman. Guidelines for the design and
statistical analysis of experiments using laboratory animals. ILAR
Journal 2005, 43, 244-258.
-
Grindstaf JL, ED Brodie & ED Ketterson. Immune function
across generations: integrating mechanism and evolutionary process
in maternal antibody transmission. Proc. Royal Society London B2003,
270, 2309-2319.
-
Hong ST, M Lee, M Seo, DH Choo, HR Moon & SH Llee.
Immunoblot analysis for serum antibodies to
Pneumocystis carinii by age and intensity
of infection in rats. Korean J. Parasitol. 1995, 33, 187-194.
-
Jonas K. Long-term holding of laboratory rodents. ILAR News
1976, 19, L1-L25.
-
Lim T-S & W-Y Loh. A comparison of tests of equality of
variances. Comp. Stat. Data Anal. 1996, 22, 287-301.
-
Manning PJ, J Gaibor, D Delong & R Gunther.
Enzyme-linked immunosorbent assay and immunoblot analysis of the
immunoglobulin G response to whole-cell and lipooligosaccharide
antigens of Pasteurella pneumotropica in laboratory mice
with latent pasteurellosis. J. Clin. Microbiol. 1989,
27, 2190-2194.
-
Motzel SL, JK Meyer & LK Riley. Detection of serum
antibodies to Bacillus piliformis in mice and rats using an
enzyme-linked immunosorbent assay. Lab. Anim. Sci. 1991, 41, 26-30.
-
Mundry R & J Fischer. Use of statistical programs for
nonparametric tests of small samples often leads to incorrect P
values: examples from animal behaviour. Animal Behav. 1998,
56, 256-259.
-
Nicklas W, M Staut & A Benner. Prevalence and
biochemical properties of V factor-dependent Pasteurellaceae from
rodents. Zentralbl. Bakteriol. 1993, 279, 114-124.
-
Nicklas W, P Baneux, R Boot, T Decelle, AA Deeny, M Fumannelli
& B Illgen-Wilke.
Recommendations for the health monitoring of rodent and rabbit
colonies in breeding and experimental units. Lab. Anim. 2002,
36, 20-42.
-
Nicklas W. Haemophilus infection in a colony of
laboratory rats. J. Clin. Microbiol. 1989, 27, 1636-1639.
-
Olsen I, FE Dewhirst, BJ Paster & HJ Busse.
Pasteurellaceae
Pp. 851-912 in: Bergey’s Manual of Systematic Bacteriology
vol. 2nd ed. Part B (Brenner DJ, Krieg NR, Staley JT eds). Springer,
New York, 2005.
Petrie A & P Watson. Statistics for Veterinary and Animal Science. Blackwell Science Ltd., London, UK 1995.
-
Siegrist CA. Mechanisms by which maternal antibodies
influence infant vaccine responses; review of hypotheses and
definition of main determinants.Vaccine 2003, 21, 3406-3412.
-
Simecka JW & GH Cassell. Serum antibody and cellular
responses in LEW end F344 rats after immunization with
Mycoplasma pulmonis antigens. Infect. Immun. 1987,
55, 731-735.
-
Suzuki E, K Miochida & M Nakagawa. Naturally occurring
subclinical Corynebacterium kutscheri infection in
laboratory rats: strain and age related antibody response. Lab.
Anim. Sci. 1988, 38, 42-45.
- Ziang ZQ & HCJ Ertl. Transfer of maternal antibodies results in inhibition of specific immune responses in the offspring. Virus Res. 1992, 24, 297-314.