
Auditory, Olfactory and Tactile Contact is not an Obstacle for Studies Involving Hormonal Interrelationships
by GA Maria1, JA Valares2, F Forcada1, & JA Abecia1*
1Departamento de Producción Animal y Ciencia de los Alimentos.
Facultad de Veterinaria, Zaragoza, Spain
2ZEU Immunotec, Zaragoza, Spain
 Correspondence: JA Abecia, PhD, Dip ECSRHM
Correspondence: JA Abecia, PhD, Dip ECSRHM
Tel +34 876 55 41 59
Fax +34 976 76 15 90
E-mail alf@unizar.es
Summary
Thirty-two ewes were used to determine whether individual housing, allowing contact with neighbours, induces a stress response. Ewes were housed in individual pens designed to allow the ewes to see, hear, smell and touch adjacent animals, and were distributed into four groups (n=8/group): ewes with subcutaneous implants containing melatonin and oestradiol (M+E), melatonin (M), oestradiol (E) and non-implanted control ewes (C). Heart rate, stress indicators (plasma cortisol, glucose, lactate and creatine kinase (CK) concentrations) and luteinizing hormone (LH) concentrations were measured hourly and compared with the resting values (before and after pen housing). Heart rate increased significantly during the introduction into the pen (P<0.001) in all groups, in comparison with the resting values. No significant differences between groups were observed for cortisol concentrations, with the exception of the M group, which showed the highest response (P<0.001) when ewes were introduced into the pens. Lactate, CK and glucose changes in comparison with the resting values were similar between groups. LH concentrations during pen housing decreased significantly in all groups in comparison with resting values. In conclusion, individual confinement of sheep allowing visual, auditory, olfactory and tactile contact with their neighbouring animals was not an obstacle for investigating particular hormonal interrelationships with multiple sampling procedures. However further investigations are required to determine if this conclusion applies to other hormone systems in sheep.
Introduction
The use of large animals as experimental models has allowed progress
in the understanding of some human physiological and pathological
mechanisms. They can provide larger volumes of sampling material
(blood, urine, faeces), and with greater frequency, than small mammals
(Arney, 2009a). Large animals have a much longer lifespan
than small mammals, which may be of interest for long term studies. In
particular, sheep (Ovis aries) are attractive animals for
medical, veterinary and fundamental biological research: they are
docile, rarely show aggression and are gregarious (Arney, 2009b). The biomedical applications of sheep as models for human diseases
have been reviewed by Scheerlinck et al.
(2008).Experimental protocols involving animal models usually
include procedures that may have the potential to cause pain or
distress to the animals. The response to stress depends on several
factors but one of the most important is the nature of the stressful
stimuli (Parrot et al., 1994). In commercial sheep management
practices, transport, manipulation, shearing or health management can
induce the stress response of the animals (Barnet & Hemsworth, 1990). These practices can also affect reproductive performance of the
ewes (Dobson et al., 2012). The nature of the stressor to
which an animal is exposed should be considered when studying the
endocrine response to adverse stimuli (Parrot et al., 1994).
A stressful environment elevates cortisol concentrations and this
could affect the pulsatility of luteinizing hormone (LH) release with
the consequent reduction of oestradiol secreted by dominant follicles,
preventing or delaying the pre-ovulatory surge (Breen & Karsch, 2004). Elevated plasma ACTH/corticosteroids concentrations have been
shown to reduce significantly the concentration of follicular LH
receptors, cause unusual pathological changes in follicles and corpora
lutea, and inhibit ovulation in ewes (López-Diaz & Bosu, 1997). This situation could negatively affect the development and
functionality of the oocyte, and the viability of the future offspring
(Dobson et al., 2012).
Individuals from gregarious domestic species can become highly
stressed if they are isolated from the social group. In sheep,
confinement and isolation cause an elevation in the cortisol
concentration which is much higher than with restraint (Parrto et al., 1994). The usual handling associated with frequent blood sampling for
hormone analysis includes spatial isolation in a pen, jugular venous
catheterization and close human contact. Some studies on sheep have
found that social isolation induced pronounced physiological stress
responses, including acceleration of the heart rate and increase in
plasma cortisol concentrations. Increased heart rate has been recorded
in relation to visual isolation of ewes (Baldock & Sibly, 1990) and restraint of the animal (Palestrini et al., 1988).
Moreover, it has been demonstrated that if one ewe is prevented from
seeing and smelling her flock mates, it causes a rise of the cortisol
concentration, which can be maintained for at least six hours (Dobson et al., 2012).
Individual housing of sheep has been a frequent practice in our
studies, especially when control of individual food consumption is
required, or a frequent bleeding regime to measure pulsatile hormones,
particularly LH, is necessary (Lozano et al., 1998; Abecia et al., 1996, 2002; Forcada et al.,
1997, 2002, 2003, 2007; Sosa et al., 2009). It is important to note that although we kept animals isolated
from their flock mates, the design of the pens used in these
experiments allowed full visual, auditory, olfactory and tactile
contact with adjacent sheep. Under these constraints, it is logical to
raise the question as to whether or not this practice could affect the
results of these studies. Treatment with melatonin has been part of
the experimental procedures of our studies, in order to determine the
effect of this hormone on LH release under different nutritional
treatments or social environments. Some authors have proposed the
hypothesis that some of the positive effects of melatonin could be
affected by a more efficient stress response of the animals treated
with this hormone (Chuang et al., 1993), or diminishing the
endocrine and behavioural impact of social isolation in ewes (Guesdon et al., 2013).
The aim of this study was to determine whether or not individual
housing which allows visual, auditory, olfactory and tactile contact
with flock mates produces a stress response, and if this response
could be affected by exogenous hormones. This is of particular
importance when hormones under study are able to modulate the
physiological adaptive syndrome per se.
Materials and methods
The study was conducted at the experimental farm of the University of Zaragoza, Spain (41°N). All procedures were approved by the in-house Ethics Committee for Animal Experiments from the University of Zaragoza (Institutional Review Board/Independent Ethics Committee number IRB00006869; Office for Human Research Protections number OHRP IORG0005699). The care and use of animals were performed according to the Spanish Policy for Animal Protection RD1201/05, which meets the European Union Directive 2010/63 on the protection of animals used for experimental and other scientific purposes.
Animals and experimental procedures
Thirty two sexually mature Rasa Aragonesa ewes were used, with a mean
(± SD) weight of 59.2±7.6 kg and a mean (± SD) body condition (score
from 0 to 5; Russel et al., 1969) of 3.10±0.47. These animals
had not been used previously for experimental purposes. Animals were
ovariectomized in the first week of August under deep anaesthesia at
the Veterinary Hospital of the University of Zaragoza. Ewes were
housed in an uncovered communal pen without supplementary light, and
always in total absence of males. In mid-October, 16 ewes (8 from the
melatonin treated group and 8 non-treated with melatonin ewes)
received a subcutaneous silastic implant (length: 1.5 cm; internal
diameter: 3.3 mm; external diameter: 4.6 mm) (Karsch et al., 1973) containing crystalline oestradiol (Sigma-Aldrich Química S.A.,
Madrid, Spain). To prevent an initial peak of steroid release,
implants were pre-soaked in water. One week after oestradiol
implantation, 16 ewes received a single subcutaneous implant
containing 18 mg melatonin (Melovine®, CEVA Salud Animal, Barcelona,
Spain) (eight of them had been previously implanted with oestradiol).
These implants were designed to maintain high plasma melatonin
concentrations for at least 90 days. Thus, animals were distributed
into four groups: ewes implanted with melatonin and oestradiol (group
M+E, n=8), ewes implanted with melatonin (group M, n=8), ewes
implanted with oestradiol (group E, n=8) and non-implanted control
ewes (group C, n=8).
On the 10th December, we housed the ewes in individual pens (2 x 2 m).
Wall pens (height 1.5 m) were made with 5 iron bars (length 2 m), so
that ewes could see, hear, smell and touch their adjacent sheep
(between 3 and 5 depending on cage’s position). Pens were
elevated on a slatted floor with automatic cleaning of manure. They
were provided with individual food and water bowls. Heart rate, stress
indicators (plasma cortisol, glucose, lactate and creatine kinase (CK)
concentrations) and plasma LH concentrations were measured at seven
occasions through the experimental period: 1) in the communal pen,
resting before uploading animals to the pens (Rest Before), 2) just
when they were introduced into the pens (Ascent), 3) after 1 hour in
the pen (Pen 1 h), 4) after 2 hours in the pen (Pen 2 h), 5) after 3
hours in the pen (Pen 3 h), 6) after 4 hours in the pen (Pen 4 h), and
7) resting 1 hour after returning the ewes to the communal pen (Rest
After). Blood samples were obtained by jugular venous catheters, which
were inserted the day before sampling by the same trained team.
Catheters were provided with a 3-way stopcock with one male luer-lock
port and two female luer-lock ports, so that heparinized saline
prevented coagulation of the catheters. Local anaesthesia was used for
the catheterization procedures. Ewes were uploaded one by one to their
pens, and immediately, the first blood sample was collected from the
first ewe. After that, the second ewe was uploaded and sampled,
followed by the other animals in the same order as before. This
stratified procedure was followed throughout the whole sampling
period. Identically, at the end of the penned period, the first ewe
was downloaded to the group pen, and sampled, then the second ewe and
so on until the last animal. Plasma was separated by centrifugation
and stored at –20° C until analysis.
Heart rate monitoring
Heart rate (beats per minute) was recorded using a Polar Sport Tester
monitor (Polar S610 tm, Polar Electro Oy, Finland), which was placed
onto each animal the day before monitoring. The transmitter was
attached to a girth belt supplied by the manufacturer for use in
humans (model S-160) and adapted to sheep with a neoprene strip. One
electrode was placed behind the scapula and the other electrode was
situated on the ventral abdomen. The receiver (codified for each ewe)
was attached to the belt on the back of the animal. To improve the
reception of the signal, the electrodes were impregnated with
ultrasound gel. The heart rate signal was telemetrically transmitted
within a range of 1 m to the receiver. The monitor calculated heart
rate based on a pulse to pulse time-averaging algorithm at 5, 15 or 60
sec intervals (Seaward et al., 1990). In this particular
study the signal was recorded every 5 sec. Data were downloaded to a
computer at the end of the study.
Hormonal and metabolite assays
Plasma glucose (mmol/L) and CK (IU/L) concentrations were determined
with a Multichannel Technicon Analyser (RA-500), using reagents for RA
Technicon systems (Bayer Diagnostics, Spain) (glucose, Ref.
T01-1492-56; CK Ref. T01-1885-01). Plasma cortisol concentrations
(nmol/L) were determined in duplicate by a single enzyme immunoassay
(EIA) (Chacon-Perez et al., 2004). The concentration of
lactate (mmol/L) was determined in fluoride oxalate plasma using a
Sigma Diagnostic Kit (Lactate 735-10) and a spectrophotometer (Lamda
5, Perkin Elmer). Plasma LH concentrations (IU/L) were measured using
a simple sandwich EIA on 96-well polyvinyl microtiter plates (Valares
et al., 2007). The intra-assay coefficients of variation were 10, 8,
14, 5 and 7% for glucose, CK, cortisol, lactate and LH, respectively.
Statistical analysis
Data were analysed using least square techniques to determine the
influence of the fixed effects included in the model. The general
representation of the model used was: y = Xb + e, where y was an N×1
vector of records, b denoted the fixed effect in the model with the
association matrix X and e was the vector of residual effects. Data
were presented as least square means ± standard error (SE). The main
effects were the treatment, with four concentrations (M, E, M+E and C)
and the seven sampling times. After testing that the interaction
effects were not significant, they were removed from the full model.
The analysis was performed using the PROC MIXED procedure of SAS
statistical software package.
Results
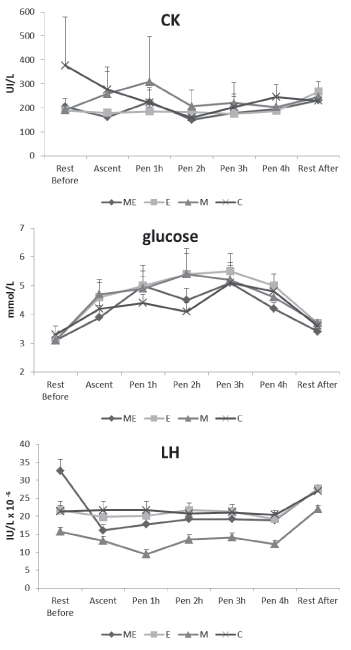
The highest mean heart rate values throughout the experiment were
observed in the E group, being significantly different to that
obtained by the other groups at most time points (P<0.001) (Figure
1). Heart rate increased significantly during the introduction to the
individual pen (P<0.001) in all the groups, in comparison with the
resting values, both before uploading to the pens and at the end of
the pen period. The relative increment of heart rate when ewes were
uploaded to the pens was lower in the M+E and E groups and higher in M
and C groups. These last two groups showed significant differences in
Pen 1h and Pen 2h time and during the rest period post pen housing
(P<0.05).
No significant differences between groups were observed for the
cortisol concentrations throughout the experiment (Figure 1), with the
exception of the M group, which showed the highest response
(P<0.001) when ewes were introduced into the pens. The variability
between groups in the cortisol concentrations was higher during the
introduction to pen (more than 1000 nmol/L) than during the end of
this stressor action (213 nmol/L).
The lactate concentrations at the beginning of the stressor action
were similar between groups, although slightly higher concentrations
were observed in the E group (Figure 1). At the end of the stressor
action the differences disappeared. Regarding CK values, no
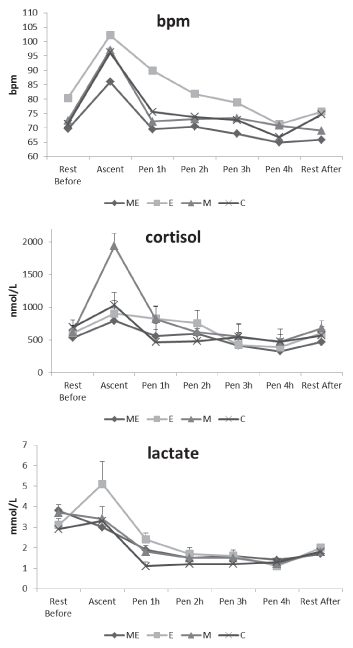
significant differences between groups were observed (Figure 2).
Introduction to the pen did not provoke any increment of plasma CK
concentrations. The glucose concentration profile during the
experiment was similar in the four groups, values increasing up to 3 h
after uploading the animals into the pens and decreasing at the end of
the experiment (Figure 2).
At the beginning of the experiment, the M+E group presented the
highest LH concentrations, being significantly different to the other
groups (P<0.001). Except in the E and C groups, occupation of the
individual pens had a reducing effect on LH concentrations, which
significantly decreased after 1 h in the pen. This reduction was shown
later, since no significant differences in the LH concentrations were
observed between hours 2, 3 and 4 compared with the 1 h concentrations
in groups, except in the M+E group. In the M group, LH release
significantly increased during the rest post pen than at the beginning
of the experiment, showing an increase of 39% in comparison with the
initial concentration.
|
|
Figure 1. Least square means (±SE) of heart rate (bpm), cortisol (nmol/L) and lactate (mmol/L) in the communal pen, resting before uploading animals to the pens (Rest Before), just when they were introduced into the pens (Ascent), after 2, 3 or 4 h in the pen (Pen 1 h, 2h, 3h, 4h) and resting 1 hour after returning the ewes to the communal pen (Rest After), of ovariectomized Rasa Aragonesa ewes treated with melatonin (M) and/or oestradiol implant (ME, E) or not treated (C).
Click image to enlarge |
|
Figure 2. Least square means (±SE) of creatine kinase (CK) (UI/L), glucose (mmol/L) and LH (IU/L) in the communal pen, resting before uploading animals to the pens (Rest Before), just when they were introduced into the pens (Ascent), after 2, 3 or 4 h in the pen (Pen 1 h, 2h, 3h, 4h) and resting 1 hour after returning the ewes to the communal pen (Rest After), of ovariectomized Rasa Aragonesa ewes treated with melatonin (M) and/or oestradiol implant (ME, E) or not treated (C).
Click image to enlarge |
Discussion
The initial rise of some the physiological stress-indicators,
coincident with the introduction to the pen and during the first
moments of isolation, demonstrates that the isolation conditions in
the present experiment induced a certain degree of stress in the short
term. The heart rate results show that the critical period was the
upload to the individual pen in all groups although it was likely due
to the exercise associated with being placed in the individual pens;
unfortunately, no behavioural indicators of stress were recorded to
confirm this observation. It has been proposed that the increased
heart rate may be a reaction to handling rather than to separation per
se (Piccione et al., 2011). Also, this could be explained by
the fact that sheep, which are a gregarious and relatively defenceless
species, show an innate and immediate response in a dangerous
situation. In this case, the initial resistance of the animal to
handling could constitute a situation of agitation corresponding to a
notable increase in heart rate (Baldock & Sibly, 1990).
In gregarious animals, behaviour among flock members is highly
synchronized. When the amount of available space increases, sheep
adjust the distance between themselves to maintain group cohesion (Sibbad et al., 2000). Extra activities are needed (i.e. locomotor) to maintain social
aggregation and hierarchy in small spaces like stalls (Piccione et al., 2011).
The lower values of heart rate at the end of the stage in the
individual pens (Pen 4 value) could be interpreted as a period of
quietness of the animals due to a combination of the protection
offered by a less variable environment (Baldock & Sibly, 1990), such as the visual, tactile and olfactory contact with social
companionsduring the stage in the pen (Gelez & Fabre-Nys, 2004), and an habituation effect to the stressor stimulus (Smith and Dobson, 2002). Thus, Hopster & Blockhuis (1994) showed that heart
rate arousal reflects the locomotor activity more than a stress
reaction per se when isolated cows are free to move. In
relation to management and human presence, some studies have shown
that sheep can habituate themselves more easily to the presence of
people than to a particular situation. In addition, the animals used
in this study had been housed on the experimental farm for at least
six months before the experiment, and have had repeated tactile,
visual and auditory contact with humans. In this context, it was
reported (Hargreaves & Hutson, 1990) that this permanent
human contact reduced the heart rate response of sheep in relation to
an approaching human.
The E group showed higher heart rates than the other groups throughout
the experiment. An increase of 15-29% in heart rate in 17-ß oestradiol
treated ovariectomized ewes compared to an untreated group has been
reported (Evans et al., 1988). In the same way, a
1 µg/kg intravenous treatment of oestradiol has been associated
with increased cardiac output and heart rate in ovariectomized ewes
(Magness & Rosenfeld, 1989). The lower heart rate
presented by the M+E group in comparison with the E group could
indicate that melatonin mitigates the stressor effect of isolation on
heart activity or the heart stimulation produced by oestradiol itself.
In fact, it has been demonstrated that melatonin decreases heart rate
(Hussein et al., 2007) and blood pressure (Koziróg et al., 2011) in men, and the administration of melatonin in rats produces a
dose-related fall in mean arterial pressure and heart rate (Chuang et al., 1993).
No significant differences between groups were observed for the
cortisol concentrations, with the exception of the M group, which
showed the significantly highest response at introduction to the pen.
An elevated adrenocortical response recorded in the serum
corticosteroid concentrations 15 h after an intraperitoneal injection
of 100 µg of melatonin in male rats has been reported (Weidenfeld et al., 1993), indicating that this hormone could directly affect plasma cortisol
concentrations. This effect was not observed in the M+E group,
indicating that the presence of exogenous oestradiol may neutralize
the potential effect of melatonin on the adaptation process.
Tilbrook et al. (2000) observed a decrease in LH
pulse frequency and amplitude during an isolation/restraint stressor
in ovariectomized oestrogen implanted ewes during the breeding season.
In our study, in ovariectomized ewes without oestrogen implants no
differences in LH pulse concentrations were observed. However, the
comparison of the current results to more frequent sampling periods,
which are required to investigate LH pulsatility and amplitude, should
be carefully considered. A decrease in the LH release (pulse frequency
and amplitude), during 4 h of transport in ovariectomized ewes with or
without prior steroid exposure at mid-breeding season has been
reported (Dobson et al., 1999). Rivier & Rivest
(1991) suggested that increased concentrations of circulating
corticosteroids do not represent the sole modulator of stress-induced
inhibition of LH secretion. On the other hand, cortisol suppresses
pulsatile LH secretion by inhibiting pituitary responsiveness to GnRH,
rather than by suppressing hypothalamic GnRH release in the
ovariectomized ewe (Breen & Karsch, 2004). It is likely
that the initial rise of cortisol concentrations observed at
introduction to the pen could be responsible for the plasma LH
decrease in the M group. However, the prolongation of stressor
stimulus does not lead to continued suppression of LH release (Smith et al., 2003). This could explain the LH results in E, M and C groups, which had
no significant differences in pen 2, pen 3 and pen 4 in comparison
with the initial concentration. Rasmussen & Malven (1983)
described that habituation to an acute confinement stress produced no
change in average plasma LH concentrations in ovariectomized ewes.
Thus, episodic secretion of LH was inhibited by the stress of initial
confinement, but several days or hours (in this case) of habituation
to the same periods of confinement minimized this inhibition and
restored the episodic discharges of LH. Moreover, an increase of mean
LH concentration after stressor stimulus (transport) has been
observed, in comparison with the values following a stressor in
ovariectomized ewes with no steroid treatment during the breeding
season (Dobson et al., 1999). These results are in accordance
with those obtained in the present study in the M and C groups.
In conclusion, individual confinement of sheep allowing visual,
auditory, olfactory and tactile contact with their neighbouring
animals was not an obstacle for investigating particular hormonal
interrelationships with multiple sampling procedures. However further
investigations are required to determine if this conclusion applies to
other hormone systems in sheep.
References
-
Abecia JA, SM Rhind, PJ Goddard, SR McMillen, S Ahmadi, & DA
Elston: Jugular and ovarian venous profiles of progesterone and
associated endometrial progesterone concentrations in pregnant and
non-pregnant ewes. Anim. Sci. 1996, 63, 229-234.
-
Abecia JA, F Forcada & O Zuniga: A note on the effect
of individual housing conditions on LH secretion in ewes after
exposure to a ram. Appl. Anim. Behav. Sci. 2002,
75, 347-352.
-
Arney DR: Welfare of large animals in scientific research.
Scand. J. Lab. Anim. Sci. 2009a, 36, 97-101.
-
Arney DR: Sheep behaviour, needs, housing and care. Scand.
J. Lab. Anim. Sci. 2009b, 36, 69-73.
-
Baldock NM & RM Sibly: Effects of handling and
transportation on the heart rate and behaviour of sheep. Appl. Anim.
Behav. Sci. 1990, 28,15-39.
-
Barnett JL & PH Hemsworth: The validity of
physiological and behavioural measures of animal welfare. Appl.
Anim. Behav. Sci. 1990, 25, 177-187.
-
Breen KM, & FJ Karsch: Does Cortisol inhibit pulsatile
luteinizing hormone secretion at the hypothalamic or pituitary
level? Endocrinology 2004, 145, 692-698.
-
Chacon-Perez G, S García-Belenguer, JC Illera & J
Palacio:
Validation of an EIA technique for the determination of salivary
cortisol in cattle. Spanish J. Agric. Res. 2004, 2,
45-51.
-
Chuang JI, SS Chen & MT Lin: Melatonin decreases brain
serotonin release, arterial pressure and heart rate in rats.
Pharmacology 1993, 47, 91–97.
-
Dobson H, JE Tebble, M Ozturk & RF Smith: Effect of
transport on pulsatile LH release in ovariectomized ewes with or
without prior steroid exposure at different times of year. J.
Reprod. Fertil. 1999, 117, 213-222.
-
Dobson H, C Fergani, JE Routly & RF Smith: Effects of
stress on reproduction in ewes. Anim. Reprod. Sci. 2012,
130, 135–140.
-
Evans W, TM Phernetton & RR Magness: 17beta-estradiol
effect on critical cardiac output with reduction of cardiac output
in oophorectomized sheep. Am. J. Physiol. 1988, 275,
57-64.
-
Forcada F, JM Lozano, JA Abecia & L Zarazaga: Control
of luteinizing hormone secretion in ewes by endogenous opioids and
the dopaminergic system during short seasonal anoestrus: role of
plane of nutrition. Anim. Sci. 1997, 65, 217-224.
-
Forcada F, O Zuniga & JA Abecia: The role of nutrition
in the regulation of LH secretion during anestrus by the
serotoninergic and dopaminergic systems in Mediterranean ewes
treated with melatonin. Theriogenology 2002, 58,
1303-1313.
-
Forcada F, JA Abecia & O Zuniga: Regulation of LH
secretion during seasonal anestrus by dopaminergic pathways in Rasa
Aragonesa ewes treated or not with melatonin. Can. J. Anim. Sci.
2003, 83, 311-313.
-
Forcada F, JA Abecia, A Casao, JA Cebrian-Perez, T Muino-Blanco
& I Palacin: Effects of ageing and exogenous melatonin on pituitary
responsiveness to GnRH in ewes during anestrus and the reproductive
season. Theriogenology 2007, 67, 855-862.
-
Gelez H & C Fabre-Nys: The “male effect” in
sheep and goats: a review of the respective roles of the two
olfactory systems. Horm. Behav. 2004, 46, 257-71.
-
Guesdon V, B Malpaux, P Delagrange, M Spedding, F Cornilleau, D
Chesneau, J Haller & E Chaillou:
Rapid effects of melatonin on hormonal and behavioral stressful
responses in ewes Psychoneuroendocrinology 2013, 38,
1426-1434.
-
Hargreaves AL & GD Hutson: The effect of gentling on
heart rate, flight distance and aversion of sheep to a handling
procedure. Appl. Anim. Behav. Sci. 1990, 26, 243-252.
-
Hopster H & HJ Blockhuis: Validation of a heart-rate
monitor for measuring a stress response in dairy cows. Can. J. Anim.
Sci. 1994, 74, 465-474.
-
Hussein MR, OG Ahmed, AF Hassan & MA Ahmed: Intake of
melatonin is associated with amelioration of physiological changes,
both metabolic and morphological pathologies associated with
obesity: an animal model. Int. J. Exp. Pathol. 2007, 88,
19-29.
-
Karsch FJ, DJ Dierschke, RF Weick, T Yamanii, J Hotchkiss & E
Knobil:
Positive and negative feedback control by estrogen of luteinizing
hormone secretion in the rhesus monkey. Endocrinology 1973,
92, 799–804.
-
Koziróg M, AR Poliwczak, P Duchnowicz, M Koter-Michalak, J
Sikora, & M Broncel:
Melatonin treatment improves blood pressure, lipid profile, and
parameters of oxidative stress in patients with metabolic syndrome.
J. Pineal Res. 2011, 50, 261–266.
-
Lopez-Diaz MC & WT Bosu: Effects of ACTH on luteinizing
hormone receptors in ovine follicular wall and corpus luteum.
Reprod. Nutri. Develop. 1997, 37, 599-612.
-
Lozano JM, Forcada F, & Abecia JA: Opioidergic and
nutritional involvement in the control of luteinizing hormone
secretion of postpartum Rasa Aragonesa ewes lambing in the
mid-breeding season. Anim. Reprod. Sci. 1998, 52,
267-277.
-
Magness RR, & CR Rosenfeld: Local and systemic
estradiol-17ß: effects on uterine and systemic vasodilation. Am.
J. Physiol. 1989, 256, 536-542.
-
Palestrini C, V Ferrante, S Mattiello, E Canali & C
Carenzi: Relationship between behaviour and heart rate as an indicator of
stress in domestic sheep under different housing systems. Small
Rumin. Res. 1988, 27, 177-181.
-
Parrott RF, HB Misson & FC de la Riva: Differential
stressor effects on the concentrations of cortisol, prolactin and
catecholamines in the blood of sheep. Res. Vet. Sci. 1994,
56, 234–239.
-
Piccione G, C Giannetto, S Marafioti, S Casella, A Assenza &
F Fazio Effect of different farming management on daily total locomotor
activity in sheep. J. Vet. Behav.: Clin. Appl. Res. 2011,
6, 243-247.
-
Rasmussen DD & PV Malven: Effects of confinement stress
on episodic secretion of LH in ovariectomized sheep.
Neuroendocrinology 1983, 36:392-396.
-
Rivier C & S Rivest: Effect of stress on the activity
of the hypothalamic-pituitary-gonadal axis: Peripheral and central
mechanisms. Biol. Reprod. 1991, 45, 523-532.
-
Russel AJF, JM Doney & RG Gunn: Subjective assessment
of body fat in live sheep. J. Agric. Sci. 1969, 72,
451-454.
-
Scheerlinck JP, KJ Snibson, VM Bowles & P Sutton:
Biomedical applications of sheep models: from asthma to vaccines.
Trends Biotechnol. 2008, 26, 259-266.
-
Seaward BI, RH Sleamaker, T McAuliffe & JF Clapp: The
precision and accuracy of a portable heart rate monitor. Biomed.
Instr. Techn. 1990, 24, 37-41.
-
Sibbad AM, LJ Shellard & TS Smart: Effects of space
allowance on the organizing behaviour and spacing of sheep. Appl.
Anim. Behav. Sci. 2000, 70, 49-62.
-
Smith RF, SP Ghuman, NP Evans, FJ Karsch & H Dobson:
Stress and the control of LH secretion in the ewe. Reproduction
2003, 61, 267-282.
-
Smith, RF & H Dobson: Hormonal interactions within the
hypothalamus and pituitary with respect to stress and reproduction
in sheep. Dom. Anim. Endocrin. 2002, 23, 75-85.
-
Sosa C, JA Abecia, M Carriquiry, F Forcada, GB Martin, I Palacin
& A Meikle:
Early pregnancy alters the metabolic responses to restricted
nutrition in sheep. Domest. Anim. Endocrin. 2009, 36,
13-23.
-
Tilbrook AJ, AI Turner & IJ Clarke: Effects of stress
on reproduction in non-rodent mammals: the role of glucocorticoids
and sex differences. Rev. Reprod. 2000, 5, 105–113.
-
Valares JA, JA Abecia, F Forcada, I Palacin, L Mata & P
Razquin:
Development of a simple enzyme immunoassay for the determination of
ovine luteinizing hormone. Vet. Res. Commun. 2007, 31,
427–436.
- Weidenfeld Y, U Schmidt, & I Nir: The effect of exogenous melatonin on the hypothalamic-pituitary-adrenal axis in intact and pinealectomized rats under basal and stressed conditions. J. Pineal. Res. 1993, 14, 60-66.