
Original scientific article
Carbonated Drinks Impact Follicle Development, Expression of Ovarian FSHR and Serum Caspase-3 in Mice
by SC Wei1, ZD Gong2*, HN Lu1, HQ Liang1, LJ Lai1
1Life Science and Engineering College, Northwest University for
Nationalities, Lanzhou, 730030, China
2Affiliated Hospital of Medicine College, Northwest
University for Nationalities, Lanzhou, 730030, China
 Correspondence: Professor Gong Zhuandi
Correspondence: Professor Gong Zhuandi
Affiliated Hospital of Medicine College, Northwest University for
Nationalities
Lanzhou, 730030, China
Tel: +0086-931-2937773
E-mail: yxgzd578@163.com
Summary
Objectives: The present study aimed to assess the
effects of Coca-Cola and Pepsi-Cola on the development of ovaries and
follicles, and on the reproduction of animals.
Methods: 150 female non-cycling mice weighing
20.36±2.28g were divided into five groups (n=30). COC-1 and COC-2 mice
drank freely 50% and 100% Coca-Cola for up to 25 days. PEP-1 and PEP-2
mice received orally 50% and 100% Pepsi-Cola. Mice in the control
group (CG) drank tap water. Another five mice were used as a normal
control per group on day 0. Ovarian and follicle indices were measured
under the microscope. Western blotting was utilized to ascertain the
expression levels of FSHR protein in ovaries. Serum concentrations of
caspase-3, epidermal growth factor (EGF) and vascular endothelial
growth factor receptor (VEGFR) were measured by ELISA.
Results: The ovarian weight of PEP-2 was
significantly lower than that of CG (P<0.05) on
day 25. Ovarian cortex thickness (OCT) of PEP-2 was significantly
reduced in comparison with CG (P<0.01). In the
COC-2 group, values of follicle longitudinal diameter (FLD), follicle
transverse diameter (FTD), follicle wall thickness (FWT), oocyte
longitudinal diameter (OLD) and oocyte transverse diameter (OTD) were
less than those of CG on day 25. FWT and OLD of PEP-1 were reduced as
compared to CG. In COC and PEP groups, numbers of POF and MF
were reduced slightly. Follicles developed poorly. FSHR protein was
decreased slightly and EGF levels increased in Coca-Cola treated or
Pepsi-Cola treated mice. Serum caspase-3 levels of COC-1 and PEP-1
were significantly lower than those of CG.
Conclusion: Coca-Cola and Pepsi-Cola treatment could
reduce ovarian weights, inhibit the ovarian cortex thickness and
impair the development of follicles.
Introduction
Carbonated beverages are the most popular types of beverage worldwide. Many people drink these almost every day (Hatch et al., 2012). However, the harmful effects of carbonated beverages have been associated with their increased consumption. The rising prevalence of overweight people, obesity and type-II diabetes or the metabolic syndrome is related to carbonated beverages (Cuomo et al., 2014; Guan et al., 2013). Recent studies have shown that consuming ≥ 1.5 servings of sugar-sweetened beverage daily could result in girls between 9 and 18.5 years old menstruating on average of 2.7 months earlier than those consuming carbonated beverages ≤ 2 times per week (Carwile et al., 2015). Consumption of carbonated drinks during menstruation could alter menstrual characteristics in nulliparous females (Adienbo et al., 2016).
It has already been demonstrated that carbonated beverages affect
adversely reproductive behaviours and functions. Ovary weights and
numbers of primary and secondary follicles were significantly reduced
in rats (Dorostghoal et al., 2011). Diameters of secondary
and antral follicles were also decreased significantly. Body weights
of Sprague–Dawley rats were increased after they were exposed to
Coca-Cola from 30, 39, or 55 weeks of age (Belpoggi et al., 2006).
Thus, carbonated beverages had noticeable effects on reproductive
endocrinology, reproductive function and fertility in women and
animals. However, most of these results were acquired based on
questionnaire investigations, or the prospective analyses of clinical
cases or epidemiological analyses, which means that the study results
are susceptible to bias and are difficult to compare (Chavarro et al., 2009). Currently, little information exists concerning the quantitative
effects of carbonated drinks on fertility (Hatch et al., 2012). Comparative experimental studies are few (Chavarro et al., 2009; Cuomo et al., 2014).
In order to examine thoroughly the influence of carbonated beverages
on the reproductive endocrinology and functions in humans, the current
investigation assessed the effects of different doses of Coca-Cola and
Pepsi-Cola on the development of ovaries and follicles of mice. The
impact of both drinks on expression levels of ovarian FSHR, and on
serum concentrations of caspase-3, epidermal growth factor (EGF) and
vascular endothelial growth factor receptor (VEGFR) was investigated.
The aim was to provide the experimental basis for further mechanistic
studies of the effects of carbonated beverages on reproductive
functions in humans.
Methods
Animals and Ethics Statement
In order to carry out a sufficiently large experiment and acquire
meaningful results, 150 non-cycling Kunming mice
(Mus musculus), 28 days old and body weight of 20.4 4±2.45g,
were purchased from the Experiment Animal Center, Lanzhou University
[License No. SCXK (Gansu) 2005-0007]. All mice were randomly assigned
into five groups (n=30): Coca-Cola group 1 (COC-1), Coca-Cola group 2
(COC-2), Pepsi-Cola group 1 (PEP-1), Pepsi-Cola group 2 (PEP-2) and
control groups (CG). All mice were accurately weighed each day using
an electronic balance, and group housed in mouse cages equipped with
automatic water dispensers in a room maintained at 22-24o and 30% to
50% relative humidity. The light cycle in the room provided 12h
light/day. Mice freely received a commercial diet (Lanzhou Taihua Feed
Co. Ltd, Lanzhou, China). Water was provided ad libitum. The
experiment was started following a 7-day adjustment period. All
animal-treatment procedures were approved by the Experiment Animal
Care and Use Committee of Gansu province, the People’s Republic
of China. All mice were treated according to humanitarian and ethical
rules.
Animal treatments and sample collection
Mice in COC-1, COC-2, PEP-1, PEP-2 and CG drank Coca-Cola, Pepsi-Cola
or tap water as outlined in Table 1. Coca-Cola and Pepsi-Cola in 5
litre bottles were bought from a supermarket in Lanzhou city. They
were stored at a room temperature of 22± 3oC. 5 mice were randomly
chosen from each group on days 5, 10, 15, 20 and 25. Each mouse was
anesthetized by injecting 0.1mg/kg xylazine intramuscularly.
Subsequently every animal was sacrificed by cervical dislocation.
Bilateral ovaries were harvested aseptically and accurately weighed
using an electronic balance. Blood samples were taken aseptically
using vacutainers (Zhejiang Gongdong Medical Technology Co. Ltd,
Zhejiang, China). Blood samples were allowed to coagulate during 2 h
at room temperature, and then were centrifuged (3000×g, 20 min). The
serum was stored at -20o until analysis.
The remaining five mice of each group and 2 healthy male mice (45 days
old) were raised together on day 25 for a week so as to mate randomly.
Additionally, another five mice per group were used as normal controls
on day 0. Bilateral ovaries and blood samples were collected on day 0
as described above.
Table 1. Administration doses of carbonated beverages and sampling. 50% Coca-Cola (Pepsi-Cola) represents one litre (1L) of pure Coca-Cola (Pepsi-Cola) diluted with the same volume of tap water (1L); 100% Coca-Cola (Pepsi-Cola) represents pure Coca-Cola (Pepsi-Cola).
| Group |
Numbers |
Treatment |
Sampling |
|
COC-1 |
30 |
50% Coca-Cola |
Ovaries and blood were collected at days 5, 10, 15, 20 and 25 |
Measurements of ovarian weight and ovarian cortex thickness
(OCT)
Each ovary was weighed using an electronic balance. Under an optical
microscope, ovarian cortex thickness (OCT) was determined immediately
with a vernier calliper. The average ovarian weight
of each mouse was determined on the basis of the right and left
values.
Histological observations and image measurement of ovaries
Ovary tissues fixed in 10% formaldehyde were sectioned (5μm), and
stained with hematoxylin and eosin (H&E). The sections were
observed under a light microscope (Leica, Japan). Microscopic images
of the ovaries were photographed. Six sites in each section (5
sections in every group, totalling 150 sites for each group) were
assessed. Ovaries and follicles were measured utilizing Images
Advanced 3.2 and Image Pro-Plus 2.0 (MOTIC Company, Hong Kong, China).
The indices included the follicle (secondary
follicles and mature follicles) longitudinal diameter
(FLD), follicle transverse diameter (FTD), follicle wall thickness
(FWT), oocyte (including the secondary oocytes and mature oocytes)
longitudinal diameter (OLD) and oocyte transverse diameter (OTD).
Western blotting analysis of FSHR protein in ovaries
To evaluate the FSHR (follicle-stimulating hormone receptor) protein
expression of ovaries following Coca-Cola and Pepsi-Cola treatment,
western blotting was conducted. Briefly, ovary samples were lysed in
lysis buffer, and the resulting proteins were subjected to 10% sodium
dodecyl sulphate-polyacrylamide gel electrophoresis, before being
transferred to polyvinylidene fluoride membranes (PVDF). These
membranes were then blocked for 2 h in a 5% non-fat milk solution
containing 10 mM Tris (pH 7.5), 100 mM NaCl, and 0.1% Tween 20 (w/v).
Membranes were incubated with rabbit anti-sheep, FSHR (Sigma, diluted
1:200), and rabbit anti-sheep β-actin (1:1000) polyclonal antibodies
at 4°C overnight, before being exposed to the appropriate secondary
antibody (1:2000) for 1 h. Mouse anti-β-actin monoclonal antibody
(1:10 000) was used as a sample loading control. Blots were developed
using a chemiluminescent reagent (SuperSignal West Pico; Thermo
Scientific, Rockford, IL, USA). The integrated optical densities of
bands in the scanned images were measured with Quantity One software
(Bio-Rad, Hercules, CA, USA). The protein expression of FSHR was
determined as the ratio of the grey value of the target band to that
of the β-actin band. Samples were run in triplicate. The negative
control was not incubated with the primary antibodies.
Detection of serum caspase-3, epidermal growth factor (EGF) and
vascular endothelial growth factor receptor (VEGFR)
Serum levels of caspase-3, epidermal growth factor (EGF) and vascular
endothelial growth factor receptor (VEGFR) were measured using ELISA
according to the manufacturer’s instructions (Shanghai Bangyi,
Biological Technology Co. Ltd, Shanghai, China). Samples were analysed
in triplicate. Analytical sensitivities were 0.10ng/mL (caspase-3) and
0.40pg/mL (EGF and VEGFR). The inter-assay CV was lower than 6%. The
correlation coefficient of the standard curve was 0.9986.
Pregnancy duration of maternal
mice and gender ratio of offspring
On day 25, female and male mice were raised together. The pregnancy
rate and duration were calculated. Numbers of
offspring and their gender ratio were also determined.
Statistical analyses
Data are reported as means ± standard errors of means. Statistical
analysis was performed with SPSS version 18.0 (SPSS Inc., Chicago, IL,
USA). After square root transformation of the data, all variables
complied with the assumptions of one-way analysis of variance (ANOVA).
Post-ANOVA comparisons between groups were carried out using the
contrast option under the general linear model procedure (Scheffé
test). When significant differences were identified, supplementary
Tukey’s post-hoc tests were conducted to investigate
pairwise differences. P-values <0.05 were considered significant.
|
|
Figure 1. A) Standard cage with aspen bedding
and nesting material. B) Experimental cage with AGREBE cotton
cloth. Click image to enlarge |
|
Figure 2. Cotton cloth showed significant
amount of wear after only few weeks of usage. A) new unused
cloth, B) cloth after one week of use, and C) cloth after three
weeks of use. Click image to enlarge |
|
Figure 2. Cotton cloth showed significant
amount of wear after only few weeks of usage. A) new unused
cloth, B) cloth after one week of use, and C) cloth after three
weeks of use. Click image to enlarge |
|
Figure 2. Cotton cloth showed significant
amount of wear after only few weeks of usage. A) new unused
cloth, B) cloth after one week of use, and C) cloth after three
weeks of use. Click image to enlarge |
Results
Ovarian weights of mice
As shown in Table 2, ovarian weights of all experimental group mice
were less than that of control group (CG) during most of the
experiment. On day 25, ovarian weight of PEP-2 was significantly lower
than that of CG (P<0.05). There were no significant
differences between Coca-Cola groups and Pepsi-Cola groups. The
findings indicate that oral ingestion of Coca-Cola and Pepsi-Cola for
a prolonged duration could reduce ovarian weights. Pepsi-Cola had a
stronger effect.
Table 2. Ovarian weights of mice (mean ± SEM, mg). Data on day 0 were measured from five normal control mice per group. *There was a significant difference when compared to control group (CG).
| Group |
0d |
5d |
10d |
15d |
20d |
25d |
COC-1 |
5.67±0.29 |
5.67±0.47 |
7.67±0.31 |
8.33±0.50 |
9.67±0.80 |
10.17±0.85 |
COC-2 |
5.33±0.49 |
5.98±0.60 |
7.02±0.65 |
8.18±0.70 |
9.33±0.89 |
9.80±0.96 |
PEP-1 |
5.87±0.51 |
6.13±0.73 |
7.33±0.74 |
8.07±0.72 |
9.27±0.58 |
9.73±0.85 |
PEP-2 |
5.83±0.51 |
6.07±0.63 |
7.01±0.72 |
8.07±0.74 |
9.17±0.82 |
9.37±0.93* |
CG |
5.67±0.52 |
6.20±0.71 |
7.33±0.71 |
8.67±0.60 |
9.67±0.88 |
10.83±1.03 |
Ovarian cortex thicknesses (OCT)
Data in Table 3 show that OCT values of experimental group mice were
lower than that of CG during the experiment. On day 25, OCT of PEP-2
was significantly reduced in comparison with CG (P<0.01).
On day 25, OCT of COC-2 and PEP-1 groups was also significantly
smaller than CG (P<0.05). The results demonstrate
that administration of Coca-Cola and Pepsi-Cola could reduce ovarian
cortex thickness. The effects increased with the duration of exposure.
The effect of Pepsi-Cola was slightly greater than that of Coca-Cola.
Table 3. Ovarian cortex thickness of mice (mean ± SEM, μm). Data on day 0 were measured from five normal control mice per group. * There was a significant difference (P<0.05) when compared to control group (CG). **There was a highly significant difference (P<0.01) when compared to control group (CG).
| Group |
0d |
5d |
10d |
15d |
20d |
25d |
COC-1 |
8.08±0.65 |
8.54±0.78 |
9.03±0.81 |
9.69±0.86 |
10.20±1.06 |
10.46±1.13 |
COC-2 |
7.98±0.62 |
8.31±0.91 |
8.37±1.01 |
8.87±0.81 |
9.12±0.85 |
9.55±0.88* |
PEP-1 |
8.06±0.78 |
8.12±0.79 |
8.27±0.75 |
8.96±0.81 |
9.01±1.04 |
9.31±1.09* |
PEP-2 |
8.11±0.81 |
8.25±0.82 |
8.29±1.86 |
8.21±0.79* |
8.12±0.76 |
8.31±0.91** |
CG |
8.12±0.79 |
8.89±0.92 |
9.28±0.91 |
10.32±0.98 |
11.38±1.12 |
12.60±1.23 |
Values of FLD, FTD, FWT, OLD and OTD
Results in Table 4 indicate that FLD, FWT, OLD and OTD of COC-2 were
less than that of CG (mostly P<0.05) on day 25. All five indices of PEP-2 were
significantly lower than CG (P<0.05 or
P<0.01). FWT and OLD of PEP-1 were decreased compared to
CG (P<0.05).
Table 4. FLD, FTD, FWT, OLD and OTD of mice on day 25 (mean ± SEM, μm). Data on day 0 were measured from five normal control mice per group. FLD-Follicle longitudinal diameter; FTD-Follicle transverse diameter; FWT-Follicle wall thickness; OLD-Oocyte longitudinal diameter; OTD-Oocyte transverse diameter. *There was a significant difference (P<0.05) when compared to control group (CG). **There was a highly significant difference (P<0.01) when compared to control group (CG).
|
Group |
FLD |
FTD |
FWT |
OLD |
OTD |
COC-1 |
407.3±39.5 |
273.3±20. 9 |
73.2±9.5 |
138.2±15.6* |
127.1±12.3 |
COC-2 |
378.5±40.7* |
251.2±21. 2 |
65.4±7.6* |
130.5±14.8* |
117.3±3.5* |
PEP-1 |
391.1±40.5 |
269.6±18.2 |
70.2±9.3* |
137.7±16.6* |
124.9±7.7 |
PEP-2 |
373.2±31.4* |
242.0±19.1* |
61.8±8.7** |
130.7±15.6* |
119.9±1.6* |
CG |
425.5±69.4 |
292.6±27.5 |
86.3±9.5 |
161.6±17.3 |
146.8±5.2 |
Histology of ovaries
The primordial follicles (POF) and primary follicles (PF) were small.
A few mature follicles (MF) existed. The structures of the ovaries and
follicles were normal. Ovarian cortex and zona pellucida (ZP) were
clear.
In COC-1 group, numbers of POF and MF were reduced slightly in
comparison with CG. A thin granular layer was distributed over the
secondary follicles (SF). In COC-2 group, numbers
of POF and PF were decreased in comparison with CG.
Small SF and MF were present. Follicles did not develop fully.
In PEP-1, histological changes were very similar to COC groups. POF
were scarcer than in COC-2. Few SF and mature MF were observed in
comparison with CG and they were larger than in the CG. The granular
layer in SF was distributed evenly. Zona pellucida (ZP) was
small. For PEP-2 group, few SF and MF existed. PF
and SF numbers were reduced as compared to COC-2.
Follicles developed poorly. Apoptosis of granular cells was observed.
The results demonstrated that drinking Coca-Cola and Pepsi-Cola for a
prolonged period could down-regulate ovary and follicle development
and maturation. Pepsi-Cola had more noticeable effects than
Coca-Cola.
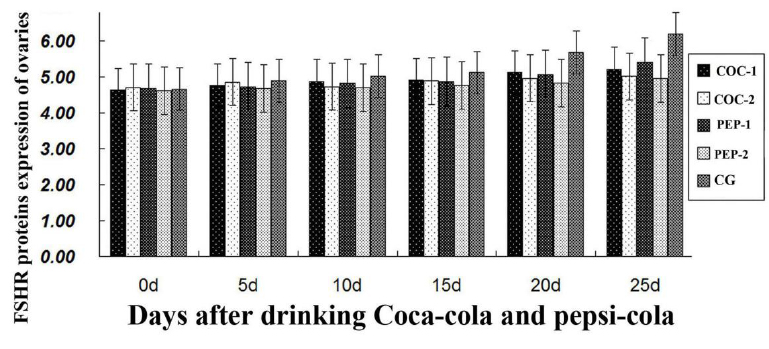
Expression levels of FSHR protein in ovaries
To assess the impact of both Coca-Cola and Pepsi-Cola treatment on
expression and mRNA level of FSHR, qRT-PCR was employed in each
group. In comparison with CG, expression level of
FSHR protein was decreased slightly in all experimental groups from
day 10 (Figure 1). However, there were no significant differences
between groups. These findings demonstrate that Coca-Cola and
Pepsi-Cola had no marked effects on the expression level of ovarian
FSHR protein in mice.
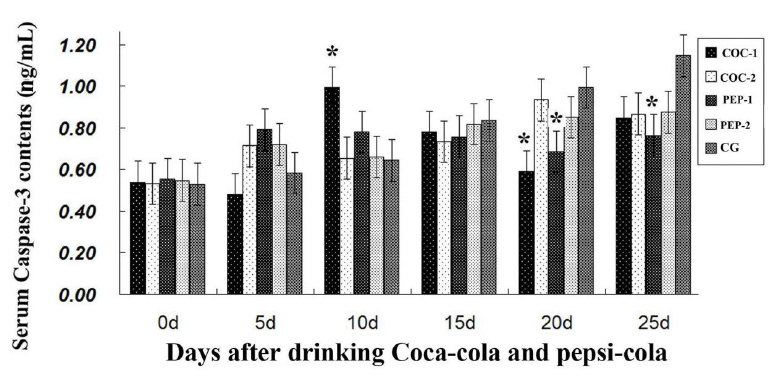
Detection of serum caspase-3
As shown in Figure 2, after day 10 serum caspase-3 levels of COC-1 and
PEP-1 decreased in comparison to CG. The maximum reduction of
caspase-3 levels was found in PEP-1 group (P<0.05). The
results indicated that Coca-Cola and Pepsi-Cola inhibited caspase-3
synthesis.
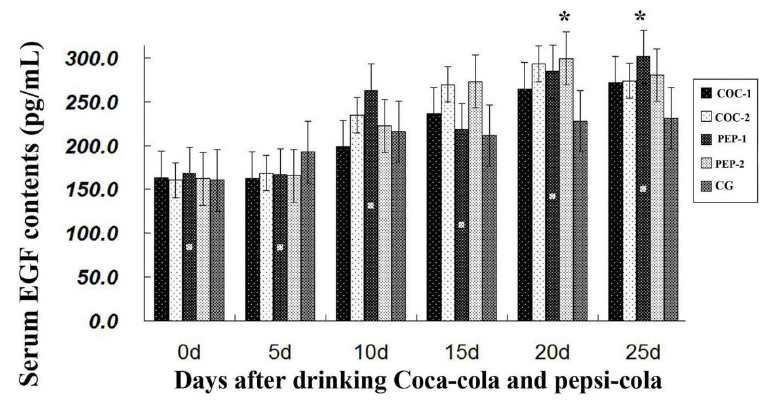
Detection of serum EGF and VEGFR
As shown in Figure 3, the epidermal growth factor (EGF) levels of the
four experimental groups were higher than that of CG after day 15. On
day 20, the level for PEP-2 was increased significantly compared to CG
(P<0.05). On day 25, the level for PEP-1
was increased significantly as compared to CG (P<0.05). Thus, Coca-Cola and Pepsi-Cola could enhance EGF
activity.
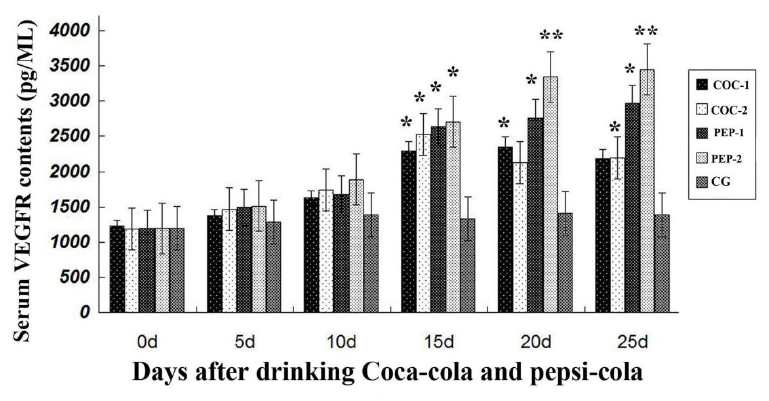
Data in Figure 4 show that serum levels of vascular endothelial growth
factor receptor (VEGFR) were increased in the four experimental
groups. From Day 15, VEGFR levels of all experimental groups were
higher than that of CG. The most significant increase was detected in
PEP-2. The results indicate that Coca-Cola and Pepsi-Cola could
increase serum VEGFR levels.
Pregnancy duration and gender ratio of offspring
Table 5 shows that there were no significant differences in pregnancy
duration between the groups. Pregnancy rate of PEP-2 mice was clearly
lower than that of CG. Mean offspring numbers were highest in COC-1
and lowest in COC-2 mice. The gender ratio (Male:Female) of offspring
decreased in experimental mice. Gender ratios of COC-1, COC-2 and
PEP-2 groups were significantly lower than that of CG (P<0.05). The survival rate and body weights of 1
week old offspring were reduced in COC-2 when compared to CG
(P<0.05). The findings demonstrate that
Coca-Cola and Pepsi-Cola clearly affected reproduction of female mice.
Table 5. Pregnancy and offspring results (mean ± SEM). Survival rate and body weights of each group were determined on day 7 after offspring birth (or 1 week old). The different capital letter superscripts represent that there was significant difference between experimental groups (P<0.05). *There was a significant difference (P<0.05) when compared to control group (CG).
|
Group |
Pregnancy rate (%) |
Pregnancy period (d) |
Offspring numbers |
Mean birth numbers |
Gender |
Body weights |
Survival |
|
COC-1 |
4 (80.0) |
20.0±0.8 |
48 |
12.0±1.1A |
40.0:60.0* |
4.05±0.41A |
46 (95.8) |
Discussion
Health concerns over carbonated soft drinks consumption have been widely reported. Many epidemiological studies have evaluated the association between caffeine and fertility, with inconsistent results. Some studies suggest that various caffeine-containing beverages may affect fertility differently (Hatch et al., 2012). Recent studies reported that caffeine (including coffee, cocoa, colas) has been associated with alterations in the levels of estradiol and other hormones in women (Kotsopoulos et al., 2009). However, the mechanisms by which caffeinated beverages affect fertility remain undetermined. Ovary weights of rats were decreased significantly in a caffeine-treated group at all stages of postnatal development (Dorostghoal et al., 2011). The number of primary and secondary follicles was decreased on days 7, 14 and 28 after birth in caffeine-treated Sprague–Dawley rats. In the present mouse study, ovarian weights of Coco-Cola and Pepsi-Cola groups were decreased compared to the control group (CG). Ovarian cortex thickness (OCT) values of the four experimental groups were decreased during the experiment in comparison with CG; the effect was most pronounced in PEP-2. FLD, FWT, OLD and OTD of COC-2 and PEP-2 were lower than that of CG on day 25. Additionally, numbers of POF and MF were slightly reduced. A thin granular layer was distributed over the secondary follicles (SF). Follicle development was inadequate. Our findings demonstrated that Coca-Cola and Pepsi-Cola affected the development of ovaries and follicles in mice. These findings were in agreement with early reports (Chavarro et al., 2009; Dorostghoal et al., 2011). But, they were in disagreement with the results of other studies (Chavarro et al., 2009; Hatch et al., 2012). Therefore, the effects and mechanism of Coca-Cola and Pepsi-Cola on ovarian and follicle development need to be thoroughly explored.
Follicle stimulating hormone (FSH) can promote the proliferation and
differentiation of preantral follicles, and thus induce follicular
growth and maturation of ovarian follicles (Miró & Hillier, 1996). The ovarian response to FSH stimulation depends on the FSHR
genotype (Perez et al., 2000; Ali et al., 2012). It has
previously been demonstrated that FSHR is expressed in multiple
ovarian cell types, including pre-ovulation granule cells and
luteinized cells. However, it was still unknown whether long-term
drinking of Coca-Cola or Pepsi-Cola influences FSHR expression in
ovaries and follicles (Gormack et al., 2015). Our study
indicates that the expression level of ovarian FSHR protein was
decreased slightly in the Coca-Cola treated and Pepsi-Cola treated
mice. To date, little information on these effects has been reported
(Imai et al., 2010; Zeng et al., 2014). Our results
remain to be confirmed by future studies.
Apoptosis is characterized by specific structural changes. Although
multiple genes are involved in apoptosis (Li et al., 2013),
key mediators are aspartate specific cysteine proteases (Caspase).
Caspase families play important roles in the process of apoptosis.
Caspase-3 is the most critical apoptosis protease in the downstream of
the caspase cascade (Cao et al., 2014). FSH down-regulates
caspase-3 mRNA level in the granule cells of dominant follicles. As a
result, FSH prevented atresia of dominant follicles (Li et al., 2013). In our present investigation, serum caspase-3 levels of COC-1 and
PEP-1 decreased with the largest effect in the PEP-1 group. The
mechanism needs to be further studied.
The epidermal growth factor (EGF) enhances epidermal regeneration,
cell motility and proliferation, and stimulates cellular migration,
proliferation and angiogenesis. The protective effect of EGF against
apoptosis is known to occur through the activation of PI3K/AKT (Zhang et al., 2014). Vascular endothelial growth factor (VEGF) and its receptor 2
(VEGFR-2) are the main promoters of angiogenesis and cellular
protection during follicular and corpus luteum (CL) development (Shibuya, 2013). VEFG directly suppressed T-cell activation via VEGFR-2 (Färkkilä et al., 2011). The main functions of VEGF and its receptors
are to control the formation of new blood vessels, and also to protect
the endothelial and granulose cells (Gavalas et al., 2012).
Vascular changes are important to regulate the follicular and CL
development and ovulation (Torres & Sanchez, 2012). It is
currently known that VEGFR can be expressed by a variety of cells (Johnson & Wilgus, 2013; Friedrich et al., 2015). Our findings indicate that serum VEGF levels of Coca-Cola treated
and Pepsi-Cola treated groups were higher than that of CG; the highest
increase was in PEP-2 mice from day 15 onwards. On day 25, EGF level
of PEP-1 was significantly increased.
Administration of Coca-Cola or Pepsi-Cola could reduce foetus numbers
and increase the proportion of female foetuses. This may be associated
with changes in the uterine micro- environment after prolonged
consumption of Coco-Cola or Pepsi-Cola. Our results need to be
confirmed in other animals and humans.
Conclusion
Oral ingestion of Coca-Cola or Pepsi-Cola over a prolonged period reduced ovarian weights, inhibited ovarian cortex thickness, and affected the development of follicles and oocytes. Coca-Cola and Pepsi-Cola reduced serum levels of caspase-3, and increased EGF and VEGFR. They clearly affected reproduction of female mice, promoted growth of offspring, and reduced pregnancy rate and affected foetus numbers. Our study has laid a solid foundation and provided the experimental data for further investigating the effects and mechanism of Coca-Cola and Pepsi-Cola on development and reproduction in humans.
Competing Interest
None of the authors has any potential financial conflict of interest related to this manuscript.
Authors Contribution
Professor Wei Suocheng was responsible for the experimental designs and writing the manuscript. Professor Gong Zhuandi raised the experimental animals and took the samples. Dr Lu Huining detected the receptor gene expressions. Miss Liang Haoqin observed the histology structures and examined the ovarian parameters. Miss Lai Luju did the statistical analyses. All authors interpreted the measures serum indices and data, critically revised the manuscript and approved the final version.
Acknowledgement
The work received the support of the National Natural Science Foundation of The People’s Republic of China (Grant No. 31460684), Changjiang Scholars and Innovative Research Team in University (IRT13091). The authors thank Dr. Martin Rooney for English revision. Manuscript English was checked using Ginger tools online.
References
- Adienbo OM, VO Hart & AA Ajah. Assessment of menstrual characteristics among nulliparous female consumers of carbonated soft drinks. Int. J. Reprod. Contracept. Obstet. Gynecol. 2016, 5(1), 194-197.
- Ali MS, MAM Khandoker, MA Afroz & AKFH Bhuiyan. Ovarian Response to Different DoseLevels of Follicle Stimulating Hormone (FSH) in Different Genotypes of Bangladeshi Cattle. Asian Australas. J. Anim. Sci. 2012, 25(1),52-8.
- Belpoggi F, M Soffritti, E Tibaldi, L Falcionil, L Bua & F Trabucco. Results of long-term carcinogenicity bioassays on Coca-Cola administered to Sprague-Dawley rats. Ann. N. Y. Acad. Sci. 2006, 1076, 736-752.
- Cao J, G Zhou, Y Liu, G Liao, Q Zhang, K Ye, D Pan & C Ou. Activation of caspase-9 and its influencing factors in beef during conditioning. Animal. 2014, 8(3), 504-509.
-
Carwile JL, WC Willett, D Spiegelman, E Hertzmark, J
Rich-Edwards, AL Frazier
& KB Michels. Sugar-sweetened beverage consumption and
age at menarche in a prospective study of US girls. Hum. Reprod.
2015, 30(3), 675-83.
-
Chavarro JE, JW Rich-Edwards, BA Rosner & WC Willett.
Caffeinated and alcoholic beverage intake in relation to ovulatory
disorder infertility. Epidemiology. 2009, 20(3),
374–381.
-
Cuomo R, P Andreozzi & FP Zito. Alcoholic beverages and
carbonated soft drinks, consumption and gastrointestinal cancer
risks. Cancer Treat. Res. 2014, 159, 97-120.
-
Dorostghoal M, MK Mahabadi & S Adham. Effects of
maternal caffeine consumption on ovarian follicle development in
Wistar rats offspring. J. Reprod. Infertil. 2011, 12(1),
15-22.
-
Friedrich RE, C Beer, M Glatzel & C Hagel. Vascular
endothelial growth factor, basic fibroblast growth factor and
epithelial growth factor receptor in peripheral nerve sheath tumors
of neurofibromatosis type 1. Anticancer. Res. 2015, 35(1),
137-44.
-
Färkkilä A, M Anttonen, J Pociuviene, A Leminen, R Butzow, M
Heikinheimo & L Unila-Kallio. Vascular endothelial growth factor (VEGF) and its receptor
VEGFR-2 are highly expressed in ovarian granulosa cell tumors. Eur.
J. Endocrinol. 2011, 164(1), 115-122.
-
Gavalas NG, M Tsiatas, O Tsitsilonis, E Politi, K Ioannou, AC
Ziogas, A Rodolakis, G Vlahos, N Thomakos, D Haidopoulos, E
Terpos, A Antsaklis, MA Dimopoulos & A Bamias. VEGF directly suppresses activation of T cells from ascites
secondary to ovarian cancer via VEGF receptor type 2. Br. J. Cancer. 2012, 107(11), 1869-1875.
-
Gormack AA, JC Peek, JG Derraik, PD Gluckman, NL Young & WS
Cutfield. Many women undergoing fertility treatment make poor lifestyle
choices that may affect treatment outcome. Hum. Reprod. 2015, 30(7), 1617–24.
-
Guan S, Y Tian & F Dong. Effect of beverage intake on
body mass index (BMI) in children and adolescents. Chinese J.
Public. Health. 2013, 29(4), 557-559.
-
Hatch EE, LA Wise, EM Mikkelsen, T Christensen, AH Riis, HT
Sørensen & KJ Rothman. Caffeinated Beverage and Soda Consumption and Time to pregnancy.
Epidemiology. 2012, 23(3), 393-401.
-
Imai A, S Ichigo, H Takagi, K Matsunami, N Suzuki & A
Yamamoto. Effects of cola intake on fertility: a review. Health. 2010,
2(9), 997-1001.
-
Johnson KE & TA Wilgus. Vascular endothelial growth
factor and angiogenesis in the regulation of cutaneous wound repair.
Adv. Wound Care. 2013, 3(10), 647-661.
-
Kotsopoulos J, AH Eliassen, SA Missmer, SE Hankinson & SS
Tworoger. Relationship between caffeine intake and plasma sex hormone
concentrations in premenopausal and postmenopausal women. Cancer.
2009, 115(12), 2765–74.
-
Li P, W Yue, Y Pang, X Yu & Y Huang. Effects of FSH and
insulin on sheep ovarian follicular granulose cells in vitro
culture. Acta Veterinaria et Zootechnica Sinica. 2013,
44(9), 1386-1391.
-
Miró F & SG Hillier. Modulation of granulosa cell
deoxyribonucleic acid synthesis and differentiation by activin.
Endocrinology. 1996, 137(2), 464-468.
-
Perez MM, J Gromoll, HM Behre, C Gassner, E Nieschlag & M
Simoni. Ovarian response to follicle-stimulating hormone
(FSH) stimulation depends on the FSH receptor genotype. J. Clin.
Endocrinol. Metabol. 2000, 85(9), 3365-3369.
-
Shibuya M. Vascular endothelial growth factor and its
receptor system, physiological functions in angiogenesis and
pathological roles in various diseases. J. Biochem. 2013,
153(1), 13-19.
-
Torres AMR & AG Sanchez. Role of vascular endothelial
growth factor (VEGF) and its receptors during the ovarian cycle.
Review. Revista Mexicana De
Ciencias Pecuaris. 2012, 3(1), 89-111.
-
Zeng H, WQ Shu, JA Chen, L Liu, DH Wang, WJ Fu, LQ Wang, JH Luo,
L Zhang, Y Tan, ZQ Qiu & YJ Huang.
Experimental comparison of the reproductive outcomes and early
development of the offspring of rats given five common types of
drinking water. Plos One. 2014, 9(10).
doi:10.1371/journal.pone.0108955.
- Zhang YF, J Pu, J Yu, M Sun, N Zhang, XQ Cao, XZ Feng, MM Ma, XY Pei & W Yanrong. EGF promotes AKT expression and phosphorylation in mouse ovarian granulosa cells through the activation of PI3K/AKT. J. Ningxia Medic. Univ. 2014, 36(5), 483-486.